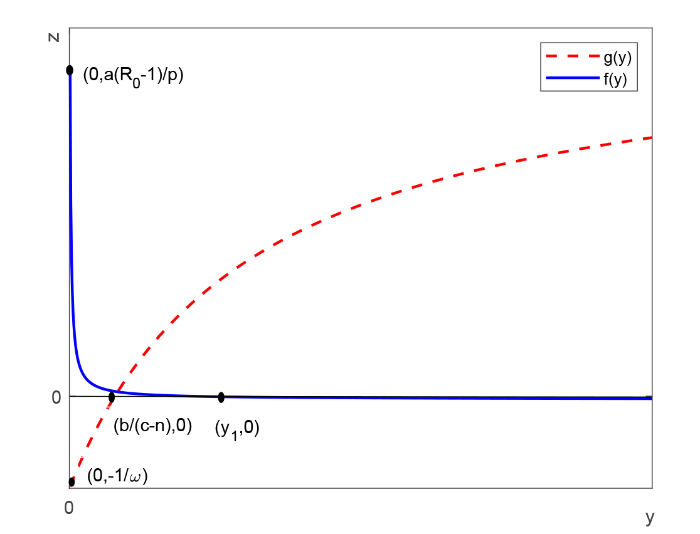
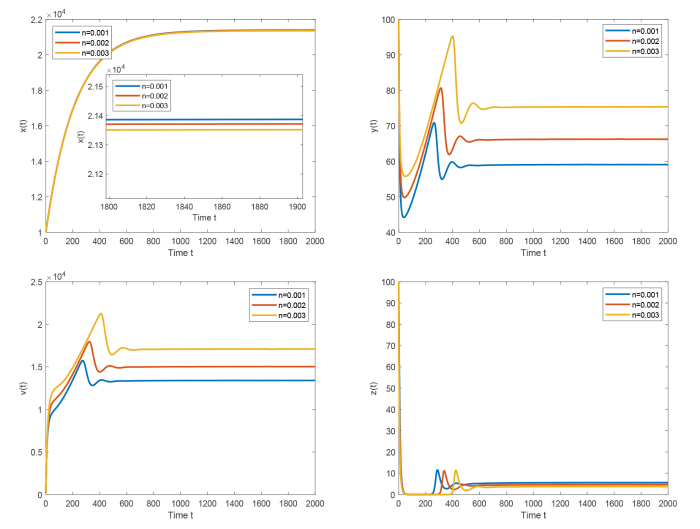
In this paper, we consider an HIV-1 infection model with cell-to-cell transmission, intracellular delay, saturated CTL immune response and immune impairment. By calculation, we get immunity-inactivated and immunity-activated reproduction ratios. By analyzing the characteristic equations, the local stability of each of feasible equilibria is established. By means of suitable Lyapunov functional and LaSalle's invariance principle, it is proved that the global asymptotic stability of each of feasible equilibria is determined by immunity-inactivated and immunity-activated reproduction ratios: If the immunity-inactivated reproduction ratio is less than unity, the infection-free equilibrium is globally asymptotically stable; if the immunity-inactivated reproduction ratio is greater than unity and the immunity-activated reproduction ratio is less than unity, the immunity-inactivated infection equilibrium is globally asymptotically stable; if the immunity-activated reproduction ratio is greater than unity, the chronic infection equilibrium is globally asymptotically stable. In addition, numerical simulation is carried out to illustrate the effects of immune impairment and cell-to-cell transmission on dynamics of the model.
Xu Rui, Zhou Kaijuan, Bai Ning. A with-in Host HIV-1 Infection Dynamics Model Based on Virus-to-cell Infection and Cell-to-cell Transmission[J]. Acta Mathematica Scientia, 2024, 44(3): 771-782
A new mathematical model was used to analyze a detailed set of human immunodeficiency virus-type 1 (HIV-1) viral load data collected from five infected individuals after the administration of a potent inhibitor of HIV-1 protease. Productively infected cells were estimated to have, on average, a life-span of 2.2 days (half-life t 1/2 = 1.6 days), and plasma virions were estimated to have a mean life-span of 0.3 days (t 1/2 = 0.24 days). The estimated average total HIV-1 production was 10.3 x 10(9) virions per day, which is substantially greater than previous minimum estimates. The results also suggest that the minimum duration of the HIV-1 life cycle in vivo is 1.2 days on average, and that the average HIV-1 generation time--defined as the time from release of a virion until it infects another cell and causes the release of a new generation of viral particles--is 2.6 days. These findings on viral dynamics provide not only a kinetic picture of HIV-1 pathogenesis, but also theoretical principles to guide the development of treatment strategies.
EggerM, HirschelB, FrancioliP, et al.
Impact of new antiretroviral combination therapies in HIV infected patients in Switzerland: Prospective multicentre study
The British Medical Journal, 1997, 315(7117): 1194-1199
A mathematical model of HIV infection: Simulating T4, T8, macrophages, antibody, and virus via specific anti-HIV response in the presence of adaptation and tropism
Bulletin of Mathematical Biology, 2010, 72(5): 1208-1253
A mathematical model of the host's immune response to HIV infection is proposed. The model represents the dynamics of 13 subsets of T cells (HIV-specific and nonspecific, healthy and infected, T4 and T8 cells), infected macrophages, neutralizing antibodies, and virus. The results of simulation are in agreement with published data regarding T4 cell concentration and viral load, and exhibit the typical features of HIV infection, i.e. double viral peaks in the acute stage, sero conversion, inverted T cell ratio, establishment of set points, steady state, and decline into AIDS. This result is achieved by taking into account thymic aging, viral and infected cell stimulation of specific immune cells, background nonspecific antigens, infected cell proliferation, viral production by infected macrophages and T cells, tropism, viral, and immune adaptation. Starting from this paradigm, changes in the parameter values simulate observed differences in individual outcomes, and predict different scenarios, which can suggest new directions in therapy. In particular, large parameter changes highlight the potentially critical role of both very vigorous and extremely damped specific immune response, and of the elimination of virus release by macrophages. Finally, the time courses of virus, antibody and T cells production and removal are systematically investigated, and a comparison of T4 and T8 cell dynamics in a healthy and in a HIV infected host is offered.
WeustenJ, DrimmelenH, LelieP N.
Mathematic modeling of the risk of HBV, HCV, and HIV transmission by window phase donations not detected by NAT
We present two HIV models that include the CTL immune response, antiretroviral therapy and a full logistic growth term for uninfected CD4+ T-cells. The difference between the two models lies in the inclusion or omission of a loss term in the free virus equation. We obtain critical conditions for the existence of one, two or three steady states, and analyze the stability of these steady states. Through numerical simulation we find substantial differences in the reproduction numbers and the behaviour at the infected steady state between the two models, for certain parameter sets. We explore the effect of varying the combination drug efficacy on model behaviour, and the possibility of reconstituting the CTL immune response through antiretroviral therapy. Furthermore, we employ Latin hypercube sampling to investigate the existence of multiple infected equilibria.
KangC J, MiaoH, ChenX, et al.
Global stability of a diffusive and delayed virus dynamics model with Crowley-Martin incidence function and CTL immune response
Advances in Difference Equations, 2017, 2017(1): 1-16
The initial stages of animal virus infection are generally described as the binding of free virions to permissive target cells followed by entry and replication. Although this route of infection is undoubtedly important, many viruses that are pathogenic for humans, including HIV-1, herpes simplex virus and measles, can also move between cells without diffusing through the extracellular environment. Cell-to-cell spread not only facilitates rapid viral dissemination, but may also promote immune evasion and influence disease. This Review discusses the various mechanisms by which viruses move directly between cells and the implications of this for viral dissemination and pathogenesis.
SattentauQ.
The direct passage of animal viruses between cells
We have developed a rapid-turnover culture system where the life span of a human immunodeficiency virus type 1-infected cell is controlled by periodic addition of a cytotoxic agent, mitomycin C. These mitomycin C-exposed cells are cocultured with a constant number of uninfected cells as new targets for the virus. Passage of the virus-infected cells under these conditions led to the emergence of a viral variant that was able to replicate efficiently in this culture system. After biologic and molecular cloning, we were able to identify a single frameshift mutation in the vpu open reading frame that was sufficient for growth of the mutant virus in the rapid-turnover assay. This virus variant spread more efficiently by cell-to-cell transfer than the parental virus did. Electron micrographs of cells infected with the delta vpu virus revealed a large number of mature viral capsids attached to the plasma membrane. The presence of these mature virus particles on the cell surface led to enhanced fusion and formation of giant syncytia with uninfected cells. Enhanced cell-to-cell transfer of the delta vpu virus provides an explanation for the survival of this mutant virus in the rapid-turnover culture system. The in vitro rapid-turnover culture system is a good representation of the in vivo turnover kinetics of infected cells and their continual replacement by host lymphopoietic mechanisms.
LaiX L, ZouX F.
Modeling cell-to-cell spread of HIV-1 with logistic target cell growth
Journal of Mathematical Analysis and Applications, 2015, 426(1): 563-584
Longitudinal studies of patients infected with HIV-1 reveal a long and variable length of asymptomatic phase between infection and development of AIDS. Some HIV infected patients are still asymptomatic after 15 or more years of infection but some patients develop AIDS within 2 years. The mechanistic basis of the disease progression has remained obscure but many researchers have been trying to explain it. For example, the possible importance of viral diversity for the disease progression and the development of AIDS has been very well worked out in the early-1990s, especially by some important works of Martin A. Nowak. These studies can give an elegant explanation for a variability of asymptomatic phase. Here, a simple mathematical model was used to propose a new explanation for a variable length of asymptomatic phase. The main idea is that the immune impairment rate increases over the HIV infection. Our model suggested the existence of so-called "Risky threshold" and "Immunodeficiency threshold" on the impairment rate. The former implies that immune system may collapse when the impairment rate of HIV exceeds the threshold value. The latter implies that immune system always collapses when the impairment rate exceeds the value. We found that the length of asymptomatic phase is determined stochastically between these threshold values depending on the virological and immunological states. Furthermore, we investigated a distribution of the length of asymptomatic phase and a survival rate of the immune responses in one HIV patient.
ElaiwA M, RaezahA A, AzozS A.
Stability of delayed HIV dynamics models with two latent reservoirs and immune impairment
Advances in Difference Equations, 2018, Atricle number 414
A mathematical model of HIV infection: Simulating T4, T8, macrophages, antibody, and virus via specific anti-HIV response in the presence of adaptation and tropism





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}