Advances in mechanisms of genetic instability related to hereditary neurological diseases
1
2005
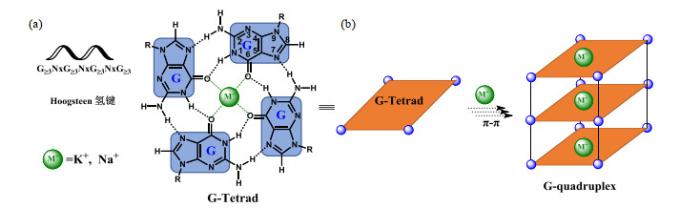
... 脱氧核糖核酸(DNA)是重要的遗传物质,不仅可以形成规整的B型双螺旋结构,还可以形成十余种不规整的非B型DNA结构,如Z-DNA、DNA错配、三螺旋(triplex)、四螺旋(quadruplex)、发卡式结构等[1 ] .G-四链体(G4)是由Gellert等[2 ] 于1962年发现的一种不规整的核酸二级结构,主要由富含鸟嘌呤(G)的DNA或RNA序列在一价阳离子(K+ 、Na+ 或NH4 + 等)的作用下,通过Hoogsteen氢键形成G-四集体(G-tetrad)(图 1 ),并进一步堆积而成. ...
Helix formation by guanylic acid
1
1962
... 脱氧核糖核酸(DNA)是重要的遗传物质,不仅可以形成规整的B型双螺旋结构,还可以形成十余种不规整的非B型DNA结构,如Z-DNA、DNA错配、三螺旋(triplex)、四螺旋(quadruplex)、发卡式结构等[1 ] .G-四链体(G4)是由Gellert等[2 ] 于1962年发现的一种不规整的核酸二级结构,主要由富含鸟嘌呤(G)的DNA或RNA序列在一价阳离子(K+ 、Na+ 或NH4 + 等)的作用下,通过Hoogsteen氢键形成G-四集体(G-tetrad)(图 1 ),并进一步堆积而成. ...
NMR spectroscopy of G-quadruplexes
2
2012
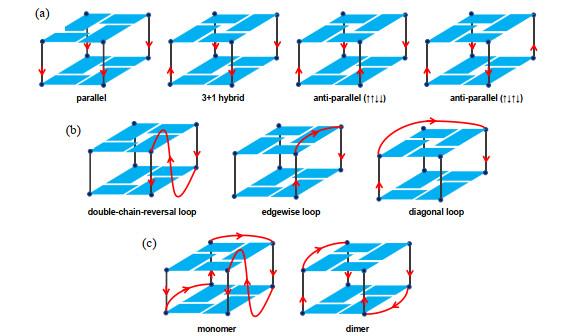
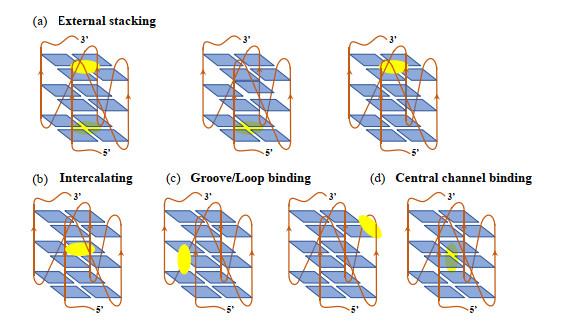
... G-四链体结构形式多样,包括:(1)链排列方向多样性.四条链有三种组合方式:四条链都平行、三条链平行与另一条链反平行、两条链平行与另外两条反平行[
图 2(a) ];(2)连接两条链的loop几何结构多样性.当G-四链体为分子内或者双分子结构时,用以连接链的loop的不同跨越会导致其多样性.比如在双分子G-四链体结构中,loop可以连接相邻或对角的链,两个loop可以头-尾或头-头方式排列[
图 2(b) ];(3)链数量多样性.形成G-四链体同一寡核苷酸可以分子内折叠,也可以分子间折叠,形成单体、二聚体或者四聚体G-四链体[
图 2(c) ].
图2 G-四链体结构多样性.(a) G-四集体的四种链朝向;(b)三种主要的Loop连接方式;(c)单体分子内和双分子分子间G-四链体.红色箭头代表链从5’到3’朝向<sup>[<xref ref-type="bibr" rid="b3">3</xref>]</sup> Structural diversity of G4 structures. (a) Four orientation of chains in G-tetrads; (b) Main three connection modes of loops; (c) Monomer or dimer G4 structure. The red arrow indicates the orientation from 5'-end to 3'-end<sup>[<xref ref-type="bibr" rid="b3">3</xref>]</sup> Fig.2 ![]()
真核和原核生物基因组都含有大量G-四链体结构.有研究[4 ] 表明,人类基因组含有约37万条可以形成G-四链体的基因序列,分布在以下几类与基因功能密切相关的区域.(1)细胞核内基因的启动子区域,特别是致癌因子(如c-MYC 、c-MYB 、BCL-2 、RET 、VEGF 、KRAS 、HIF-1 α [5 ] ;也可在转录时产生单链DNA,影响mRNA合成. (2) 端粒区域.端粒DNA是真核生物染色体线性DNA,3’末端一段富含G重复序列,用以维持染色体的稳定.当该G链悬突中有G-四链体形成时,会影响端粒酶的逆转录活性,从而导致端粒不能正常延伸.(3)细胞质中单链mRNA区域也能形成G-四链体,使得核糖体无法识别mRNA上密码子,导致翻译停止,从而下调基因表达[6 ] .(4)外显子及内含子区域.当转录通过编码链上富含G的序列时,G-四链体的形成会导致基因组的不稳定,触发机体转录偶联修复(TCR)途径,修补基因组中具有转录活性的基因编码链上不正常结构引起的损伤.G-四链体的形成会使RNA聚合酶II发生停滞,使mRNA合成受阻.修复过程会招募TCR因子,但此过程较容易引起不必要的TCR,造成无损伤部位产生重复的修复补丁;在高转录区域,这种情况会导致自发突变水平增加[7 ] . ...
... [
3 ]
Fig.2 真核和原核生物基因组都含有大量G-四链体结构.有研究[4 ] 表明,人类基因组含有约37万条可以形成G-四链体的基因序列,分布在以下几类与基因功能密切相关的区域.(1)细胞核内基因的启动子区域,特别是致癌因子(如c-MYC 、c-MYB 、BCL-2 、RET 、VEGF 、KRAS 、HIF-1 α [5 ] ;也可在转录时产生单链DNA,影响mRNA合成. (2) 端粒区域.端粒DNA是真核生物染色体线性DNA,3’末端一段富含G重复序列,用以维持染色体的稳定.当该G链悬突中有G-四链体形成时,会影响端粒酶的逆转录活性,从而导致端粒不能正常延伸.(3)细胞质中单链mRNA区域也能形成G-四链体,使得核糖体无法识别mRNA上密码子,导致翻译停止,从而下调基因表达[6 ] .(4)外显子及内含子区域.当转录通过编码链上富含G的序列时,G-四链体的形成会导致基因组的不稳定,触发机体转录偶联修复(TCR)途径,修补基因组中具有转录活性的基因编码链上不正常结构引起的损伤.G-四链体的形成会使RNA聚合酶II发生停滞,使mRNA合成受阻.修复过程会招募TCR因子,但此过程较容易引起不必要的TCR,造成无损伤部位产生重复的修复补丁;在高转录区域,这种情况会导致自发突变水平增加[7 ] . ...
![]()
Four-stranded nucleic acids: structure, function and targeting of G-quadruplexes
1
2008
... 真核和原核生物基因组都含有大量G-四链体结构.有研究[4 ] 表明,人类基因组含有约37万条可以形成G-四链体的基因序列,分布在以下几类与基因功能密切相关的区域.(1)细胞核内基因的启动子区域,特别是致癌因子(如c-MYC 、c-MYB 、BCL-2 、RET 、VEGF 、KRAS 、HIF-1 α [5 ] ;也可在转录时产生单链DNA,影响mRNA合成. (2) 端粒区域.端粒DNA是真核生物染色体线性DNA,3’末端一段富含G重复序列,用以维持染色体的稳定.当该G链悬突中有G-四链体形成时,会影响端粒酶的逆转录活性,从而导致端粒不能正常延伸.(3)细胞质中单链mRNA区域也能形成G-四链体,使得核糖体无法识别mRNA上密码子,导致翻译停止,从而下调基因表达[6 ] .(4)外显子及内含子区域.当转录通过编码链上富含G的序列时,G-四链体的形成会导致基因组的不稳定,触发机体转录偶联修复(TCR)途径,修补基因组中具有转录活性的基因编码链上不正常结构引起的损伤.G-四链体的形成会使RNA聚合酶II发生停滞,使mRNA合成受阻.修复过程会招募TCR因子,但此过程较容易引起不必要的TCR,造成无损伤部位产生重复的修复补丁;在高转录区域,这种情况会导致自发突变水平增加[7 ] . ...
Targeting G-quadruplexes in gene promoters: a novel anticancer strategy?
1
2011
... 真核和原核生物基因组都含有大量G-四链体结构.有研究[4 ] 表明,人类基因组含有约37万条可以形成G-四链体的基因序列,分布在以下几类与基因功能密切相关的区域.(1)细胞核内基因的启动子区域,特别是致癌因子(如c-MYC 、c-MYB 、BCL-2 、RET 、VEGF 、KRAS 、HIF-1 α [5 ] ;也可在转录时产生单链DNA,影响mRNA合成. (2) 端粒区域.端粒DNA是真核生物染色体线性DNA,3’末端一段富含G重复序列,用以维持染色体的稳定.当该G链悬突中有G-四链体形成时,会影响端粒酶的逆转录活性,从而导致端粒不能正常延伸.(3)细胞质中单链mRNA区域也能形成G-四链体,使得核糖体无法识别mRNA上密码子,导致翻译停止,从而下调基因表达[6 ] .(4)外显子及内含子区域.当转录通过编码链上富含G的序列时,G-四链体的形成会导致基因组的不稳定,触发机体转录偶联修复(TCR)途径,修补基因组中具有转录活性的基因编码链上不正常结构引起的损伤.G-四链体的形成会使RNA聚合酶II发生停滞,使mRNA合成受阻.修复过程会招募TCR因子,但此过程较容易引起不必要的TCR,造成无损伤部位产生重复的修复补丁;在高转录区域,这种情况会导致自发突变水平增加[7 ] . ...
G-quadruplex structures: in vivo evidence and function
1
2009
... 真核和原核生物基因组都含有大量G-四链体结构.有研究[4 ] 表明,人类基因组含有约37万条可以形成G-四链体的基因序列,分布在以下几类与基因功能密切相关的区域.(1)细胞核内基因的启动子区域,特别是致癌因子(如c-MYC 、c-MYB 、BCL-2 、RET 、VEGF 、KRAS 、HIF-1 α [5 ] ;也可在转录时产生单链DNA,影响mRNA合成. (2) 端粒区域.端粒DNA是真核生物染色体线性DNA,3’末端一段富含G重复序列,用以维持染色体的稳定.当该G链悬突中有G-四链体形成时,会影响端粒酶的逆转录活性,从而导致端粒不能正常延伸.(3)细胞质中单链mRNA区域也能形成G-四链体,使得核糖体无法识别mRNA上密码子,导致翻译停止,从而下调基因表达[6 ] .(4)外显子及内含子区域.当转录通过编码链上富含G的序列时,G-四链体的形成会导致基因组的不稳定,触发机体转录偶联修复(TCR)途径,修补基因组中具有转录活性的基因编码链上不正常结构引起的损伤.G-四链体的形成会使RNA聚合酶II发生停滞,使mRNA合成受阻.修复过程会招募TCR因子,但此过程较容易引起不必要的TCR,造成无损伤部位产生重复的修复补丁;在高转录区域,这种情况会导致自发突变水平增加[7 ] . ...
Transcriptional processing of G4 DNA
1
2009
... 真核和原核生物基因组都含有大量G-四链体结构.有研究[4 ] 表明,人类基因组含有约37万条可以形成G-四链体的基因序列,分布在以下几类与基因功能密切相关的区域.(1)细胞核内基因的启动子区域,特别是致癌因子(如c-MYC 、c-MYB 、BCL-2 、RET 、VEGF 、KRAS 、HIF-1 α [5 ] ;也可在转录时产生单链DNA,影响mRNA合成. (2) 端粒区域.端粒DNA是真核生物染色体线性DNA,3’末端一段富含G重复序列,用以维持染色体的稳定.当该G链悬突中有G-四链体形成时,会影响端粒酶的逆转录活性,从而导致端粒不能正常延伸.(3)细胞质中单链mRNA区域也能形成G-四链体,使得核糖体无法识别mRNA上密码子,导致翻译停止,从而下调基因表达[6 ] .(4)外显子及内含子区域.当转录通过编码链上富含G的序列时,G-四链体的形成会导致基因组的不稳定,触发机体转录偶联修复(TCR)途径,修补基因组中具有转录活性的基因编码链上不正常结构引起的损伤.G-四链体的形成会使RNA聚合酶II发生停滞,使mRNA合成受阻.修复过程会招募TCR因子,但此过程较容易引起不必要的TCR,造成无损伤部位产生重复的修复补丁;在高转录区域,这种情况会导致自发突变水平增加[7 ] . ...
Site-specific oligonucleotide binding represses transcription of the human c-myc gene in vitro
1
1988
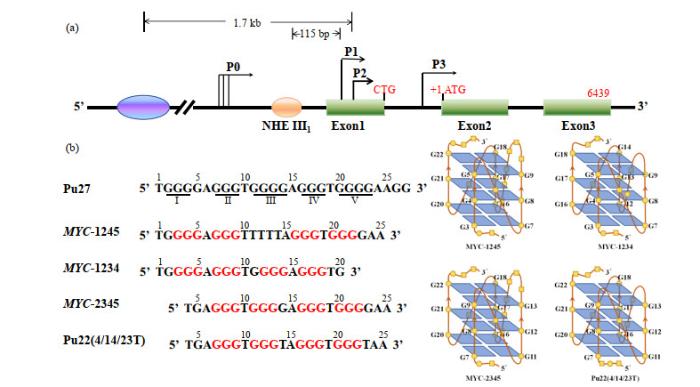
... 异常的MYC 基因在肿瘤细胞中的表达主要在转录水平被调节.c-MYC 启动子P1近端核酸酶超敏元件Ⅲ1 (NHE Ⅲ1 )控制该基因高达90%的转录激活,而NHE Ⅲ1 区域富含碱基G,可以形成G-四链体结构,对c-MYC 转录具有负调控作用,这表明NHE Ⅲ1 序列二级结构处于B型双螺旋和非B型结构动态平衡中[8 , 9 ] .天然状态下,该区域形成四种平行G-四链体结构混合体,四种结构的区别在于形成loop碱基数不同(图 3 ).当这些G-四链体与小分子配体TMPyP4相互作用时,转变成平行与反平行混合的G-四链体结构. ...
Activating transcription from single stranded DNA
1
1996
... 异常的MYC 基因在肿瘤细胞中的表达主要在转录水平被调节.c-MYC 启动子P1近端核酸酶超敏元件Ⅲ1 (NHE Ⅲ1 )控制该基因高达90%的转录激活,而NHE Ⅲ1 区域富含碱基G,可以形成G-四链体结构,对c-MYC 转录具有负调控作用,这表明NHE Ⅲ1 序列二级结构处于B型双螺旋和非B型结构动态平衡中[8 , 9 ] .天然状态下,该区域形成四种平行G-四链体结构混合体,四种结构的区别在于形成loop碱基数不同(图 3 ).当这些G-四链体与小分子配体TMPyP4相互作用时,转变成平行与反平行混合的G-四链体结构. ...
Design and synthesis of an expanded porphyrin that has selectivity for the c-MYC G-quadruplex structure
4
2005
... 当NHE Ⅲ1 富含G区域的序列处于单链状态时,转录激活因子CNBP(CCHC-type zinc finger Nucleic acid-Binding Protein)及hnRNP K(heterogeneous nuclear RiboNucleoProtein K)和单链DNA结合,促进MYC 转录进行;当该序列被TMPyP4或Se2SAP稳定时,G-四链体阻碍CNBP及hnRNP K与NHE Ⅲ1 结合,转录被抑制[10 , 11 ] (图 4 ). ...
... [
10 ,
11 ]
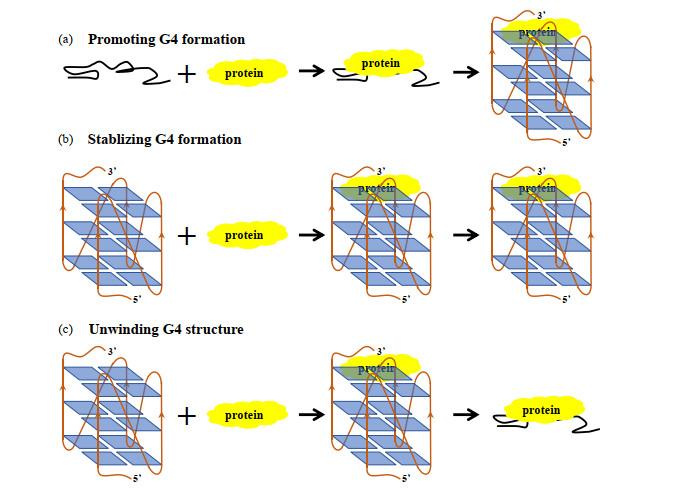
Transcriptional regulation of G-quadruplex structure formed in c-<i>MYC</i> promoter NHE Ⅲ<sub>1</sub><sup>[<xref ref-type="bibr" rid="b10">10</xref>, <xref ref-type="bibr" rid="b11">11</xref>]</sup> Fig.4 c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
![]()
... [
10 ,
11 ]
Fig.4 c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
![]()
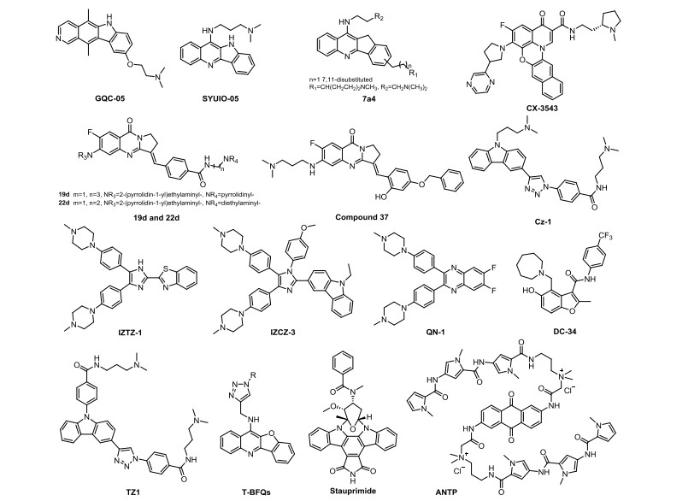
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription
3
2002
... 当NHE Ⅲ1 富含G区域的序列处于单链状态时,转录激活因子CNBP(CCHC-type zinc finger Nucleic acid-Binding Protein)及hnRNP K(heterogeneous nuclear RiboNucleoProtein K)和单链DNA结合,促进MYC 转录进行;当该序列被TMPyP4或Se2SAP稳定时,G-四链体阻碍CNBP及hnRNP K与NHE Ⅲ1 结合,转录被抑制[10 , 11 ] (图 4 ). ...
... ,
11 ]
Transcriptional regulation of G-quadruplex structure formed in c-<i>MYC</i> promoter NHE Ⅲ<sub>1</sub><sup>[<xref ref-type="bibr" rid="b10">10</xref>, <xref ref-type="bibr" rid="b11">11</xref>]</sup> Fig.4 c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
![]()
... ,
11 ]
Fig.4 c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
![]()
Developing novel G-quadruplex ligands: from interaction with nucleic acids to interfering with nucleic acid(-)protein interaction
1
2019
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
DNA and RNA quadruplex-binding proteins
2014
The regulation and functions of DNA and RNA G-quadruplexes
1
2020
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
Identification and characterization of nucleolin as a c-MYC G-quadruplex-binding protein
1
2009
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
The C-terminus of nucleolin promotes the formation of the c-MYC G-quadruplex and inhibits c-MYC promoter activity
1
2010
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
Nucleophosmin C-terminal leukemia-associated domain interacts with G-rich quadruplex forming DNA
1
2010
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
G-quadruplex DNA recognition by nucleophosmin: new insights from protein dissection
2014
Structure of nucleophosmin DNA-binding domain and analysis of its complex with a G-quadruplex sequence from the c-MYC promoter
1
2012
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
Novel interaction of the Z-DNA binding domain of human ADAR1 with the oncogenic c-MYC promoter G-quadruplex
1
2014
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
Wild-type p53 binds to MYC promoter G-quadruplex
1
2016
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
Human Pif1 helicase is a G-quadruplex DNA-binding protein with G-quadruplex DNA-unwinding activity
1
2010
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
A parallel quadruplex DNA is bound tightly but unfolded slowly by pif1 helicase
2015
Pif1 helicase unfolding of G-quadruplex DNA is highly dependent on sequence and reaction conditions
1
2018
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
NMR investigation of the interaction between the RecQ C-terminal domain of human bloom syndrome protein and G-quadruplex DNA from the human c-MYC promoter
1
2019
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
RecQ-core of BLM unfolds telomeric G-quadruplex in the absence of ATP
2014
Mechanistic insight into the interaction of BLM helicase with intra-strand G-quadruplex structures
1
2014
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
RecQ family helicases: roles in cancer and aging
1
2000
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
The Bloom's syndrome helicase unwinds G4 DNA
1
1998
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
DDX5 helicase resolves G-quadruplex and is involved in MYC gene transcriptional activation
1
2019
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
Human c-MYC transcription factor PuF identified as nm23-H2 nucleoside diphosphate kinase, a candidate suppressor of tumor metastasis
1
1993
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
The transcription factor, Nm23H2, binds to and activates the translocated c-MYC allele in Burkitt's lymphoma
1
1995
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
The guanine-quadruplex structure in the human c-myc gene's promoter is converted into B-DNA form by the human poly(ADP-ribose)polymerase-1
1
2012
... c-MYC G4-DNA本身不能参与肿瘤细胞生长与发育过程,需要在相关蛋白存在下才能发挥调节功能.如图 5 所示,这些蛋白通过与c-MYC G4-DNA结合,促进、稳定或解开其G4-DNA结构来介导MYC 基因的转录[12 -14 ] .如蛋白nucleolin(NCL)的RNA结合结构域(RBDs)3、(RBDs)4,及RGG(Arg-Gly-Gly)结构域与NHE Ⅲ1 区富含G序列作用能促进c-MYC G4-DNA形成,稳定其结构[15 , 16 ] ,抑制MYC 基因转录.CNBP蛋白是首个报道的富含G的DNA或RNA结合蛋白,能促进G4-DNA形成,致使MYC 基因转录片刻内被抑制,同时招募NM23-H2,激活MYC 基因转录.核仁磷酸蛋白NPM1(NucleoPhosMin 1)C端NPM1-C70[17 -19 ] 及作用于双链RNA 1的腺苷脱氨基化酶ADR1(Adenosine Deaminase acting on RNA 1)的Z-DNA结合域[20 ] 都能结合单链NHE Ⅲ1 富含G序列,促进c-MYC G4-DNA形成,抑制MYC 基因转录.肿瘤抑制因子P53突变体C端片段320-393 aa与c-MYC G4-DNA结合作用很强[21 ] ,可在肿瘤细胞中调节基因表达.解螺旋酶Pif1[22 -24 ] 、BLM[25 -27 ] 、WRN[28 , 29 ] 、DDX5[30 ] 、NM23-H2[31 , 32 ] 及PARP-1[33 ] 等通过与c-MYC G4-DNA作用,使其去折叠,抑制癌症转移. ...
Inhibition of cell proliferation by quindoline derivative (SYUIQ-05) through its preferential interaction with c-MYC promoter G-quadruplex
2
2011
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
... [34 -37 ]),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Solution structure of a 2:1 quindoline-c-MYC G-quadruplex: insights into G-quadruplex-interactive small molecule drug design
1
2011
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Discovery of Novel 11-triazole substituted benzofuro[3, 2-b]quinolone derivatives as c-MYC G-quadruplex specific stabilizers via click chemistry
2017
New Disubstituted quindoline derivatives inhibiting Burkitt's lymphoma cell proliferation by impeding c-MYC transcription
1
2017
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Stabilization and structure of telomeric and c-MYC region intramolecular G-quadruplexes: the role of central cations and small planar ligands
1
2007
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
The cationic porphyrin TMPyP4 down-regulates c-MYC and human telomerase reverse transcriptase expression and inhibits tumor growth in vivo
2002
The dynamic character of the G-quadruplex element in the c-MYC promoter and modification by TMPyP4
2004
Small-molecule interaction with a five-guanine-tract G-quadruplex structure from the human MYC promoter
1
2005
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Discovery of a new four-leaf clover-like ligand as a potent c-MYC transcription inhibitor specifically targeting the promoter G-quadruplex
1
2018
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
A drug-like imidazole-benzothiazole conjugate inhibits malignant melanoma by stabilizing the c-MYC G-quadruplex
1
2020
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
New substituted quinoxalines inhibit triple-negative breast cancer by specifically downregulating the c-MYC transcription
1
2019
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Carbazole ligands as c-MYC G-quadruplex binders
1
2018
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Small molecule driven stabilization of promoter G-quadruplexes and transcriptional regulation of c-MYC
1
2018
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Occurrence, biogenesis, and synthesis of biologically active carbazole alkaloids
1
2012
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
9-N-substituted berberine derivatives: stabilization of G-quadruplex DNA and down-regulation of oncogene c-MYC
1
2008
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Bisaryldiketene derivatives: A new class of selective ligands for c-myc G-quadruplex DNA
1
2010
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Design, synthesis, and evaluation of isaindigotone derivatives to downregulate c-MYC transcription via disrupting the interaction of NM23-H2 with G-quadruplex
2
2017
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
... [50 ],bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
A selective G-quadruplex DNA-stabilizing ligand based on a cyclic naphthalene diimide derivative
1
2015
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Structure-based optimization of FDA-approved drug methylene blue as a c-MYC G-quadruplex DNA stabilizer
2
2011
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
... [52 ],氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
pH-mediated fluorescence and G-quadruplex binding of amido phthalocyanines
1
2010
... 目前蛋白质结构数据库(PDB)库中与c-MYC G4相关的结构数量有197个,其中c-MYC G4与小分子复合物的结构数量为十余个.这为理解相关的分子识别机制奠定了基础.如图 7 所示,文献报道的能够结合并稳定c-MYC G4-DNA结构,并且已进入临床前或者临床II期研究的小分子药物包括:喹叨啉类(quindoline)化合物(如SYUIO-05[34 ] 、7a4[35 ] 、T-BFQs[34 -37 ] ),阳离子卟啉类(cationic porphyrins)[10 , 38 -41 ] ,咪唑类[42 , 43 ] ,喹喔啉类(quinoxalines)[44 ] ,咔唑类(carbazoles)[45 -47 ] (如Cz-1[46 ] ),小檗碱衍生物[48 -50 ] ,isaindigotone衍生物[50 ] ,bisaryldiketene衍生物[49 ] ,环萘二酰亚胺类(cyclic naphthalene diimide)衍生物[51 , 52 ] ,亚甲蓝(methylene blue)衍生物[52 ] ,氨基酞菁(amido phthalocyanine)[53 ] 等.但是,尽管这么多化合物能够靶向c-MYC G4-DNA,显示抑制MYC 基因转录的效果,但迄今为止仍然没有一个小分子化合物被批准上市,主要原因如下:(1)目前所有报道的G4-DNA、G4-RNA结构具有很高的相似性;因此设计出来的小分子有可能与其他G4-DNA或者G4-RNA相互作用,造成“脱靶”效应;(2)靶向c-MYC G4-DNA的小分子,一方面可以促进c-MYC G4-DNA结构的形成,另一方面也会促进其他G4结构的形成,从而干扰正常细胞发挥功能.因此靶向c-MYC G4-DNA的小分子药物的设计需要新的思路. ...
Circular dichroism and guanine quadruplexes
1
2012
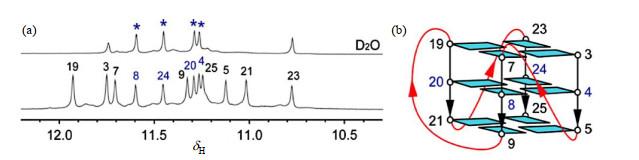
... 研究G-四链体结构常用的技术手段有:(1)圆二色谱(CD)与其他技术的联用.CD可以作为一个首选方法用来判断有无G-四链体形成[54 ] ,电泳、层析及质谱法可以给出分子的大小[55 , 56 ] .但这些方法均不能给出原子分辨率的结构信息.(2)X-ray单晶衍射技术.但是G-四链体分子在水溶液中不易结晶,因此不是首选方法.(3)NMR技术.NMR可以监测和分辨多种构象.G-四链体中鸟嘌呤的亚氨基质子,其化学位移在一般在δ 10~12范围内;而Watson-Crick碱基互补配对中亚氨基质子的化学位移则位于更低场(δ 13~14).位于中间G-四集体上的鸟嘌呤的亚氨基质子与溶剂重水交换非常慢(图 8 ),利于G-四链体拓扑结构测定. ...
Formation of parallel four-stranded complexes by guanine-rich motifs in DNA and its implications for meiosis
1
1988
... 研究G-四链体结构常用的技术手段有:(1)圆二色谱(CD)与其他技术的联用.CD可以作为一个首选方法用来判断有无G-四链体形成[54 ] ,电泳、层析及质谱法可以给出分子的大小[55 , 56 ] .但这些方法均不能给出原子分辨率的结构信息.(2)X-ray单晶衍射技术.但是G-四链体分子在水溶液中不易结晶,因此不是首选方法.(3)NMR技术.NMR可以监测和分辨多种构象.G-四链体中鸟嘌呤的亚氨基质子,其化学位移在一般在δ 10~12范围内;而Watson-Crick碱基互补配对中亚氨基质子的化学位移则位于更低场(δ 13~14).位于中间G-四集体上的鸟嘌呤的亚氨基质子与溶剂重水交换非常慢(图 8 ),利于G-四链体拓扑结构测定. ...
Sequence and solvent effects on telomeric DNA bimolecular G-quadruplex folding kinetics
1
2013
... 研究G-四链体结构常用的技术手段有:(1)圆二色谱(CD)与其他技术的联用.CD可以作为一个首选方法用来判断有无G-四链体形成[54 ] ,电泳、层析及质谱法可以给出分子的大小[55 , 56 ] .但这些方法均不能给出原子分辨率的结构信息.(2)X-ray单晶衍射技术.但是G-四链体分子在水溶液中不易结晶,因此不是首选方法.(3)NMR技术.NMR可以监测和分辨多种构象.G-四链体中鸟嘌呤的亚氨基质子,其化学位移在一般在δ 10~12范围内;而Watson-Crick碱基互补配对中亚氨基质子的化学位移则位于更低场(δ 13~14).位于中间G-四集体上的鸟嘌呤的亚氨基质子与溶剂重水交换非常慢(图 8 ),利于G-四链体拓扑结构测定. ...
Solution structure of all parallel G-quadruplex formed by the oncogene RET promoter sequence
2
2011
... 研究G-四链体结构常用的技术手段有:(1)圆二色谱(CD)与其他技术的联用.CD可以作为一个首选方法用来判断有无G-四链体形成
[54 ] ,电泳、层析及质谱法可以给出分子的大小
[55 , 56 ] .但这些方法均不能给出原子分辨率的结构信息.(2)X-ray单晶衍射技术.但是G-四链体分子在水溶液中不易结晶,因此不是首选方法.(3)NMR技术.NMR可以监测和分辨多种构象.G-四链体中鸟嘌呤的亚氨基质子,其化学位移在一般在
δ 10~12范围内;而Watson-Crick碱基互补配对中亚氨基质子的化学位移则位于更低场(
δ 13~14).位于中间G-四集体上的鸟嘌呤的亚氨基质子与溶剂重水交换非常慢(
图 8 ),利于G-四链体拓扑结构测定.
图8 (a) 三片层G-四链体在H<sub>2</sub>O(下)中及交换到D<sub>2</sub>O中1 h(上)后的亚氨基质子的一维氢谱;(b) <i>RET</i> G-四链体的拓扑结构<sup>[<xref ref-type="bibr" rid="b57">57</xref>]</sup> (a) Imino proton region of one-dimensional <sup>1</sup>H NMR spectra in H<sub>2</sub>O (down) and D<sub>2</sub>O (upper, exchanged from H<sub>2</sub>O sample 1 h later), respectively. (b) <i>RET</i> G4-DNA structure<sup>[<xref ref-type="bibr" rid="b57">57</xref>]</sup> Fig.8 ![]()
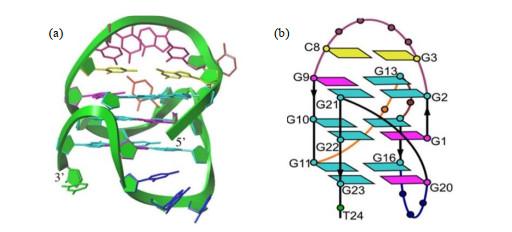
利用NMR技术可以非常方便地解析分子量比较小、不易长单晶的G4-DNA或G4-RNA与小分子配体复合物结构.有研究[58 ] 报道,VEGFR-2在肿瘤生长过程中表达上调,抑制VEGFR-2的表达可以抑制血管生成,进而阻止肿瘤生长,而且VEGFR-2还与增殖性视网膜疾病密切相关.因此VEGF和VEGFR-2可以作为潜在的抗肿瘤相关药物的靶点,阻断VEGF/VEGFR-2信号通路,或者阻止VEGF与VEGFR-2蛋白的结合均可作为抗肿瘤药物设计的着手点[58 ] .vegfr-2 基因启动子区-120到-90之间存在一段GC富集区,位于该基因的转录起始位点上游,对其转录具有调控作用.2018年,Liu等[59 ] 利用NMR技术确定了该区域碱基序列突变体VEGFR-17T的G4-DNA结构(图 9 ),它由三个叠置的G-tetrad组成,包含三个syn 鸟嘌呤.第一个鸟嘌呤G1位于中央G-四分体内.一个非典型的v形环跨越三个G-四集体平面的loop,不含有任何碱基.此外,还含有一个碱基序列较长的且呈现对角线形状的环状loop,包括六个核苷酸,连接着反向的双链. ...
... [
57 ]
Fig.8 利用NMR技术可以非常方便地解析分子量比较小、不易长单晶的G4-DNA或G4-RNA与小分子配体复合物结构.有研究[58 ] 报道,VEGFR-2在肿瘤生长过程中表达上调,抑制VEGFR-2的表达可以抑制血管生成,进而阻止肿瘤生长,而且VEGFR-2还与增殖性视网膜疾病密切相关.因此VEGF和VEGFR-2可以作为潜在的抗肿瘤相关药物的靶点,阻断VEGF/VEGFR-2信号通路,或者阻止VEGF与VEGFR-2蛋白的结合均可作为抗肿瘤药物设计的着手点[58 ] .vegfr-2 基因启动子区-120到-90之间存在一段GC富集区,位于该基因的转录起始位点上游,对其转录具有调控作用.2018年,Liu等[59 ] 利用NMR技术确定了该区域碱基序列突变体VEGFR-17T的G4-DNA结构(图 9 ),它由三个叠置的G-tetrad组成,包含三个syn 鸟嘌呤.第一个鸟嘌呤G1位于中央G-四分体内.一个非典型的v形环跨越三个G-四集体平面的loop,不含有任何碱基.此外,还含有一个碱基序列较长的且呈现对角线形状的环状loop,包括六个核苷酸,连接着反向的双链. ...
![]()
Arenobufagin, a bufadienolide compound from toad venom, inhibits VEGF-mediated angiogenesis through suppression of VEGFR-2 signaling pathway
2
2012
... 利用NMR技术可以非常方便地解析分子量比较小、不易长单晶的G4-DNA或G4-RNA与小分子配体复合物结构.有研究[58 ] 报道,VEGFR-2在肿瘤生长过程中表达上调,抑制VEGFR-2的表达可以抑制血管生成,进而阻止肿瘤生长,而且VEGFR-2还与增殖性视网膜疾病密切相关.因此VEGF和VEGFR-2可以作为潜在的抗肿瘤相关药物的靶点,阻断VEGF/VEGFR-2信号通路,或者阻止VEGF与VEGFR-2蛋白的结合均可作为抗肿瘤药物设计的着手点[58 ] .vegfr-2 基因启动子区-120到-90之间存在一段GC富集区,位于该基因的转录起始位点上游,对其转录具有调控作用.2018年,Liu等[59 ] 利用NMR技术确定了该区域碱基序列突变体VEGFR-17T的G4-DNA结构(图 9 ),它由三个叠置的G-tetrad组成,包含三个syn 鸟嘌呤.第一个鸟嘌呤G1位于中央G-四分体内.一个非典型的v形环跨越三个G-四集体平面的loop,不含有任何碱基.此外,还含有一个碱基序列较长的且呈现对角线形状的环状loop,包括六个核苷酸,连接着反向的双链. ...
... [58 ].vegfr-2 基因启动子区-120到-90之间存在一段GC富集区,位于该基因的转录起始位点上游,对其转录具有调控作用.2018年,Liu等[59 ] 利用NMR技术确定了该区域碱基序列突变体VEGFR-17T的G4-DNA结构(图 9 ),它由三个叠置的G-tetrad组成,包含三个syn 鸟嘌呤.第一个鸟嘌呤G1位于中央G-四分体内.一个非典型的v形环跨越三个G-四集体平面的loop,不含有任何碱基.此外,还含有一个碱基序列较长的且呈现对角线形状的环状loop,包括六个核苷酸,连接着反向的双链. ...
A putative G-quadruplex structure in the proximal promoter of VEGFR-2 has implications for drug design to inhibit tumor angiogenesis
3
2018
... 利用NMR技术可以非常方便地解析分子量比较小、不易长单晶的G4-DNA或G4-RNA与小分子配体复合物结构.有研究[58 ] 报道,VEGFR-2在肿瘤生长过程中表达上调,抑制VEGFR-2的表达可以抑制血管生成,进而阻止肿瘤生长,而且VEGFR-2还与增殖性视网膜疾病密切相关.因此VEGF和VEGFR-2可以作为潜在的抗肿瘤相关药物的靶点,阻断VEGF/VEGFR-2信号通路,或者阻止VEGF与VEGFR-2蛋白的结合均可作为抗肿瘤药物设计的着手点[58 ] .vegfr-2 基因启动子区-120到-90之间存在一段GC富集区,位于该基因的转录起始位点上游,对其转录具有调控作用.2018年,Liu等[59 ] 利用NMR技术确定了该区域碱基序列突变体VEGFR-17T的G4-DNA结构(图 9 ),它由三个叠置的G-tetrad组成,包含三个syn 鸟嘌呤.第一个鸟嘌呤G1位于中央G-四分体内.一个非典型的v形环跨越三个G-四集体平面的loop,不含有任何碱基.此外,还含有一个碱基序列较长的且呈现对角线形状的环状loop,包括六个核苷酸,连接着反向的双链. ...
... [
59 ]
Solution structure of <i>vegfr-2</i> 17T G4-DNA displayed in ribbon mode (a) and cartoon mode (b), respectively, in which <i>syn</i>-G bases are shown in purple, <i>anti</i>-G bases are shown in cyan, G-C base-pairs are shown in yellow<sup>[<xref ref-type="bibr" rid="b59">59</xref>]</sup> Fig.9 2018年,Calabrese等[60 ] 利用NMR技术解析了小分子化合物DC-34与c-MYC G4-DNA复合物的溶液三维结构(图 10 ),发现两个DC-34分子以满负荷的方式分别结合在c-MYC G4-DNA的5’及3’端. ...
![]()
... [
59 ]
Fig.9 2018年,Calabrese等[60 ] 利用NMR技术解析了小分子化合物DC-34与c-MYC G4-DNA复合物的溶液三维结构(图 10 ),发现两个DC-34分子以满负荷的方式分别结合在c-MYC G4-DNA的5’及3’端. ...
![]()
Chemical and structural studies provide a mechanistic basis for recognition of the MYC G-quadruplex
3
2018
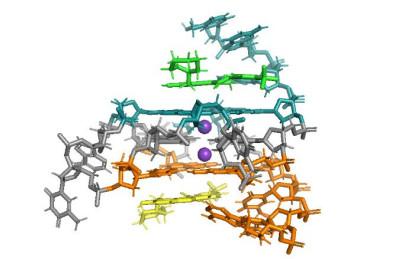
... 2018年,Calabrese等[60 ] 利用NMR技术解析了小分子化合物DC-34与c-MYC G4-DNA复合物的溶液三维结构(图 10 ),发现两个DC-34分子以满负荷的方式分别结合在c-MYC G4-DNA的5’及3’端. ...
... [
60 ]
NMR structure of DC-34 complex with c-<i>MYC</i> G4. (PDB: 5W77). All molecules are shown as sticks. Two DC-34 molecules are shown in green and yellow, and K<sup>+</sup> cations are shown as purple spheres<sup>[<xref ref-type="bibr" rid="b60">60</xref>]</sup> Fig.10 2020年,Wang等[61 ] 利用NMR技术筛选1 000多个小分子后,发现天然产物秋水仙素(colchicine)能够特异性结合肿瘤因子RET 启动子区G4-DNA结构(图 11 ).他们利用NMR技术进一步确定了colchicine结合RET G4-DNA的复合物结构[61 ] ,发现colchicine结合在RET G4-DNA的3’端,通过破坏碱基G14与RET G4-DNA原有的骨架G-tetrad之间的π-π堆积作用而与RET G4-DNA的3’端的G-tetrad发生堆积作用.这种作用方式与非特异性结合的配体berberine类似[62 ] ,并没有因为小分子结构的差异结合方式发生变化.小分子结构分析表明,导致秋水仙素能够特异性结合RET G4-DNA的原因在于,秋水仙素的小分子结构刚性不及berberine,精细的柔性结构导致其不能与c-MYC G4-DNA、Tel G4-DNA、c-Kit G4-DNA相互作用,而只与RET G4-DNA相互作用. ...
![]()
... [
60 ]
Fig.10 2020年,Wang等[61 ] 利用NMR技术筛选1 000多个小分子后,发现天然产物秋水仙素(colchicine)能够特异性结合肿瘤因子RET 启动子区G4-DNA结构(图 11 ).他们利用NMR技术进一步确定了colchicine结合RET G4-DNA的复合物结构[61 ] ,发现colchicine结合在RET G4-DNA的3’端,通过破坏碱基G14与RET G4-DNA原有的骨架G-tetrad之间的π-π堆积作用而与RET G4-DNA的3’端的G-tetrad发生堆积作用.这种作用方式与非特异性结合的配体berberine类似[62 ] ,并没有因为小分子结构的差异结合方式发生变化.小分子结构分析表明,导致秋水仙素能够特异性结合RET G4-DNA的原因在于,秋水仙素的小分子结构刚性不及berberine,精细的柔性结构导致其不能与c-MYC G4-DNA、Tel G4-DNA、c-Kit G4-DNA相互作用,而只与RET G4-DNA相互作用. ...
![]()
Colchicine selective interaction with oncogene RET G-quadruplex revealed by NMR
4
2020
... 2020年,Wang等[61 ] 利用NMR技术筛选1 000多个小分子后,发现天然产物秋水仙素(colchicine)能够特异性结合肿瘤因子RET 启动子区G4-DNA结构(图 11 ).他们利用NMR技术进一步确定了colchicine结合RET G4-DNA的复合物结构[61 ] ,发现colchicine结合在RET G4-DNA的3’端,通过破坏碱基G14与RET G4-DNA原有的骨架G-tetrad之间的π-π堆积作用而与RET G4-DNA的3’端的G-tetrad发生堆积作用.这种作用方式与非特异性结合的配体berberine类似[62 ] ,并没有因为小分子结构的差异结合方式发生变化.小分子结构分析表明,导致秋水仙素能够特异性结合RET G4-DNA的原因在于,秋水仙素的小分子结构刚性不及berberine,精细的柔性结构导致其不能与c-MYC G4-DNA、Tel G4-DNA、c-Kit G4-DNA相互作用,而只与RET G4-DNA相互作用. ...
... [61 ],发现colchicine结合在RET G4-DNA的3’端,通过破坏碱基G14与RET G4-DNA原有的骨架G-tetrad之间的π-π堆积作用而与RET G4-DNA的3’端的G-tetrad发生堆积作用.这种作用方式与非特异性结合的配体berberine类似[62 ] ,并没有因为小分子结构的差异结合方式发生变化.小分子结构分析表明,导致秋水仙素能够特异性结合RET G4-DNA的原因在于,秋水仙素的小分子结构刚性不及berberine,精细的柔性结构导致其不能与c-MYC G4-DNA、Tel G4-DNA、c-Kit G4-DNA相互作用,而只与RET G4-DNA相互作用. ...
... [
61 ]
Structure of <i>RET</i> G4-DNA in complex with colchicine (in orange). (a) The ensemble of 20 structures with the lowest energy. Base G14 is flexible; (b) One conformer of the complex. An angle 27.5° was formed between aromatic seven-membered ring and phenyl ring; (c) The structure of free <i>RET</i> G4-DNA; (d) The position of colchicine (in orange) relative to G14 (in green) above G3-G9-G13-G19 tetrad (in cyan) at 3'-end in the complex structure, compared to structure of free <i>RET</i> G4-DNA (in line mode, except the base G14 which was termed as G14 free and displayed in blue stick). In (a)~(c), the G-tetrad composed by four <i>syn</i> guanines was in magenta line and cartoon modes, respectively. The G-tetrads composed by <i>anti-</i> guanines were shown in cyan lines and cartoon modes, respectively. In all figures, base G14 was in deep-gray lines and cartoon modes. Bases G16 and T20 were in green lines and cartoon modes, respectively. Bases G4, C5, G6 and C10 in loops were in wheat lines or cartoon modes. All nonpolar protons were not displayed<sup>[<xref ref-type="bibr" rid="b61">61</xref>]</sup> Fig.11 4 总结与展望 MYC 在人类大部分癌细胞中高表达,抑制其转录是治疗肿瘤的有效手段.c-MYC 启动子区P1近端NHE Ⅲ1 区域富含碱基G序列形成G-四链体,调控c-MYC 基因转录,是抗肿瘤药物靶标.目前靶向c-MYC G4抗癌药物因为“脱靶”效应,存在很多困难.研究MYC 启动区G4-DNA的结构,有利于靶向c-MYC 基因启动子区G4-DNA的抗癌药物的开发和临床推进,从而提高癌症病人的存活率.NMR在分子量较小的G4-DNA及G4-RNA结构确定中具有冷冻电镜、X-单晶衍射不可取代的特色作用,在靶向G4-DNA小分子药物筛选方面也能发挥前期引导应用,因此,结合NMR、小分子药物有机合成和药物选择性相互作用确认等技术,靶向c-MYC G4-DNA的抗肿瘤药物的开发将会迎来新局面. ...
![]()
... [
61 ]
Fig.11 4 总结与展望 MYC 在人类大部分癌细胞中高表达,抑制其转录是治疗肿瘤的有效手段.c-MYC 启动子区P1近端NHE Ⅲ1 区域富含碱基G序列形成G-四链体,调控c-MYC 基因转录,是抗肿瘤药物靶标.目前靶向c-MYC G4抗癌药物因为“脱靶”效应,存在很多困难.研究MYC 启动区G4-DNA的结构,有利于靶向c-MYC 基因启动子区G4-DNA的抗癌药物的开发和临床推进,从而提高癌症病人的存活率.NMR在分子量较小的G4-DNA及G4-RNA结构确定中具有冷冻电镜、X-单晶衍射不可取代的特色作用,在靶向G4-DNA小分子药物筛选方面也能发挥前期引导应用,因此,结合NMR、小分子药物有机合成和药物选择性相互作用确认等技术,靶向c-MYC G4-DNA的抗肿瘤药物的开发将会迎来新局面. ...
![]()
NMR studies on the interaction between oncogene RET G-quadruplex and berberine
1
2020
... 2020年,Wang等[61 ] 利用NMR技术筛选1 000多个小分子后,发现天然产物秋水仙素(colchicine)能够特异性结合肿瘤因子RET 启动子区G4-DNA结构(图 11 ).他们利用NMR技术进一步确定了colchicine结合RET G4-DNA的复合物结构[61 ] ,发现colchicine结合在RET G4-DNA的3’端,通过破坏碱基G14与RET G4-DNA原有的骨架G-tetrad之间的π-π堆积作用而与RET G4-DNA的3’端的G-tetrad发生堆积作用.这种作用方式与非特异性结合的配体berberine类似[62 ] ,并没有因为小分子结构的差异结合方式发生变化.小分子结构分析表明,导致秋水仙素能够特异性结合RET G4-DNA的原因在于,秋水仙素的小分子结构刚性不及berberine,精细的柔性结构导致其不能与c-MYC G4-DNA、Tel G4-DNA、c-Kit G4-DNA相互作用,而只与RET G4-DNA相互作用. ...






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}