

1 引言
世界多个地方都曾出现过动物种群的炭疽大爆发, 引起了许多学者的关注和研究. 1981 年, Furniss 和 Hahn[4]以克鲁格国家公园爆发的动物炭疽为背景, 建立了差分方程炭疽模型. 2013 年, Friedman 和 Yakubu[5]在动物种群中建立了一个带有局部扩散的微分方程模型, 研究了尸体摄入、尸体引起的环境污染和迁移率等对动物种群的影响. 2017 年, Saad-Roy 等[6]通过建立具有 Logistic 增长的炭疽模型来研究动物群体中炭疽的传播, 其研究结果表明, 染病动物的数量与炭疽孢子病毒的数量密切相关. 在近几年的传染病建模中, 媒介在疾病传播过程中的作用越来越受到人们的关注. 从十九世纪开始, 昆虫就被认为是传播炭疽的罪魁祸首, 在二十世纪初,叮咬蝇被认为是疾病传播的重要媒介[7]. 以往的一些文章中指出在实验条件下至少有 21 种虻科蝇被证实其身体部位携带炭疽杆菌[8], 表明炭疽能够通过苍蝇进行传播. 2014 年, Blackburn 等[9]研究了尸食性蝇传播炭疽的方式, 文章中指出当动物感染炭疽死亡后, 由于染病尸体上寄生着寻找食物和产卵的尸食性蝇, 因此, 一方面蛆和苍蝇可能会污染尸体外部及尸体附近的土壤, 另一方面离开尸体的成年苍蝇能够通过呕吐或排便的方式将携带的细菌转移到附近植被, 进而感染食草动物. 此外, 文献[10,11]也对尸食性蝇的传播方式进行了介绍. 2013 年, Baldacchino 等[12]研究了血食性蝇传播炭疽的方式, 由于血食性蝇叮咬染病动物后嘴部或腿部携带炭疽杆菌, 则之后通过叮咬能够传染易感动物. 2010 年, Blackburn 等[13]在文章中指出了尸食性蝇和血食性蝇均能够传播炭疽. 2017 年, Mushayabasa 等[14]在建立动物种群中炭疽的传播模型时加入了血食性蝇, 研究了所建模型的理论结果与媒介对炭疽传播和控制的影响.
在研究媒介对炭疽传播的影响时, 一方面以往的大部分文章虽然介绍了动物种群与尸食性蝇、血食性蝇之间复杂的传播机制, 但是没有建立相应的模型进行分析, 另一方面一些学者只研究了一类苍蝇对动物种群中炭疽传播的影响, 不够全面, 在此基础上, 本文同时考虑了尸食性蝇和血食性蝇两类苍蝇在食草动物中的传播机制, 建立了一个具有两类媒介的动物种群中炭疽传播的数学模型.
2 模型的建立
本文考虑了十一个仓室, 分别为易感动物 (Sa(t)), 染病动物 (Ia(t)), 环境中的孢子病毒 (P(t)), 自然死亡的动物尸体 (Sc(t)), 感染炭疽死亡的动物尸体 (Ic(t)), 则动物的总数量为Na(t)=Sa(t)+Ia(t), 尸体的总数量为Nc(t)=Sc(t)+Ic(t). 对于尸食性蝇和血食性蝇两类媒介, 考虑它们的幼虫阶段和成虫阶段, 并将成虫阶段分为两类. 假设尸食性蝇 (血食性蝇) 的幼虫数量为LN(t)(LH(t)), 成虫易感者数量为SN(t)(SH(t)), 成虫染病者数量为IN(t)(IH(t)). 其具体模型为
其中, 易感动物的输入率为Λa, 自然死亡率为da,βPa是环境中孢子病毒对易感动物的传染率. 由于尸食性蝇从染病尸体上获得孢子病毒后通过污染尸体附近的植被感染食草动物, 血食性蝇通过叮咬染病动物携带孢子病毒进而感染食草动物, 则βHa,βcN和βaH分别表示染病的血食性蝇对易感动物的传染率、染病尸体对尸食性蝇的传染率和染病动物对血食性蝇的传染率. 染病动物的因病死亡率为δa,ηc是染病尸体释放孢子病毒的速率, 尸食性蝇感染炭疽后释放到尸体附近植被的孢子病毒的速率为ηN,ξ是孢子病毒的衰减率,μc是尸体的腐烂率. 对于尸食性蝇和血食性蝇两类媒介, 假设感染炭疽不影响产卵率bN(bH) 和死亡率dN(dH), 幼虫期的自然死亡率和成熟率分别为μN(μH) 和ΛN(ΛH), 在苍蝇繁殖过程中, 幼虫因拥挤或竞争而死亡的现象很常见,αN(αH) 表示幼虫的密度依赖性发育死亡率. 系统 (2.1) 中所有参数均为正常数. 为了简便, 引入变量FN(t)=SN(t)+IN(t),FH(t)=SH(t)+IH(t), 则系统 (2.1) 的等价系统为
下面证明系统 (2.2) 的有界性.
定理 2.1 对于任意的初值(Sa(0),Ia(0),P(0),Sc(0),Ic(0),LN(0),FN(0),IN(0),LH(0),FH(0),IH(0))∈Ω, 系统 (2.2) 存在唯一的非负有界解, 其中
此外, 系统 (2.2) 的正向不变集为
将系统 (2.2) 中的Sa与Ia,Sc与Ic分别相加可得
则对任意的t>0,Na(t)>0且Nc(t)>0, 此时Na(t)和Nc(t)在t∈[0,T)上有界.
对于以下系统
由文献[16]可知, 对R2+∖{(0,0)}中的所有初值, 存在一个全局渐近稳定的平衡点(bλαd,bα(λd)2). 由于
通过比较原理可知, 在[0,T)上LN(t)和FN(t)有界. 类似地,LH(t)和FH(t)在[0,T)上有界.
注意到P′(t)=ηcIc(t)+ηNIN(t)−ξP(t)≤ηcNc(t)+ηNFN(t)−ξP(t), 再次使用比较定理可得P(t)在t∈[0,T)上有界. 因此T=∞,Γ中的估计不等式可由上面的讨论得到. 定理得证.
从系统 (2.2) 可以看出尸食性蝇满足的方程为
这一系统在文献[17]中已经被研究了. 定义尸食性蝇的媒介再生数为RνN=bNΛNdN(μN+ΛN). 由系统 (2.6) 可知,当RνN≤1时, 系统(2.6)存在一个平衡点(0,0), 当RνN>1时, 系统 (2.6)存在正平衡点(L∗N,F∗N), 其中
血食性蝇满足的方程为
类似地, 定义血食性蝇的媒介再生数为RνH=bHΛHdH(μH+ΛH), 当RνH≤1时, 系统 (2.7) 存在一个平衡点(0,0), 当RνH>1时, 系统 (2.7) 存在一个正平衡点(L∗H,F∗H), 其中
根据文献[17]可知以下结论是成立的.
引理 2.1 (i) 对于系统 (2.6), 若RνN≤1, 平衡点(0,0)在R2+上是全局渐近稳定的; 若RνN>1, 正平衡点(L∗N,F∗N)在R2+∖{(0,0)}上是全局渐近稳定的.
(ii) 对于系统 (2.7), 若RνH≤1, 平衡点(0,0)在R2+上是全局渐近稳定的; 若RνH>1, 正平衡点(L∗H,F∗H)在R2+∖{(0,0)}上是全局渐近稳定的.
3 平衡点的存在性和各类再生数
下面计算系统 (2.2) 的平衡点. 显然, 系统 (2.2) 有以下五个平衡点, 分别为
(1)E1=(S0a,0,0,S0c,0,0,0,0,0,0,0),S0a=Λada,S0c=Λaμc.
平衡点E1是两类苍蝇均不存在时模型的无病平衡点, 且E1总存在.
(2)E2=(Sa2,Ia2,P2,Sc2,Ic2,0,0,0,0,0,0), Sa2=ξμcβPaηc, Ia2=daξμc(βPaηcΛaξμcda−1)βPaηc(da+δa), P2=daβPa(βPaηcΛaξμcda−1), Sc2=daξβPaηc, Ic2=daξβPaηc(βPaηcΛaξμcda−1).
平衡点E2是两类苍蝇均不存在时模型的地方病平衡点, 当且仅当βPaηcΛaξμcda>1时,E2存在.
(3)E3=(S0a,0,0,S0c,0,0,0,0,L∗H,F∗H,0),S0a=Λada,S0c=Λaμc,L∗H=ΛH+μHαH(RνH−1),F∗H=ΛHdHL∗H.
平衡点E3是只存在血食性蝇时模型的无病平衡点, 当且仅当RνH>1时,E3存在.
(4)E4=(S0a,0,0,S0c,0,L∗N,F∗N,0,0,0,0),S0a=Λada,S0c=Λaμc,L∗N=ΛN+μNαN(RνN−1),F∗N=ΛNdNL∗N.
平衡点E4是只存在尸食性蝇时模型的无病平衡点, 当且仅当RνN>1时,E4存在.
(5)E5=(S0a,0,0,S0c,0,L∗N,F∗N,0,L∗H,F∗H,0), S0a=Λada, S0c=Λaμc, L∗N=ΛN+μNαN(RνN−1), F∗N=ΛNdNL∗N, L∗H=ΛH+μHαH(RνH−1), F∗H=ΛHdHL∗H.
平衡点E5是系统 (2.2) 的无病平衡点, 当且仅当RνN>1,RνH>1时,E5存在.
其中RA为两类蝇均不存在时模型的基本再生数,RN为只存在尸食性蝇时模型的基本再生数,RH为只存在血食性蝇时模型的基本再生数,R0为两类蝇都存在时模型的基本再生数.
系统 (2.2) 的其他平衡点计算如下.
首先计算只存在尸食性蝇时模型的地方病平衡点, 令LH=FH=IH=0. 设E6=(Sa6,Ia6,P6,Sc6,Ic6,LN6,FN6,IN6,0,0,0)是系统 (2.2) 的平衡点, 由系统 (2.2) 可得:Sa6=Λa−μcIc6da, Ia6=μcIc6da+δa, Sc6=Λa−μcIc6μc, P6=ηcIc6+ηNIN6ξ, LN6=L∗N=ΛN+μNαN(RνN−1), FN6=F∗N=ΛNdNL∗N, IN6=βcNF∗NIc6βcNIc6+dNS0c. 仅考虑Ic6≠0的情况, 则关于Ic6的方程为
其中B1=βPaβcNμcηcda>0,B2=βcNξμc(1−RA)+βPaμcda(ηcdNΛaμc+ηNβcNF∗N),B3=dNΛaξ(1−RA−βPaηNβcNF∗NξdNda). 易知B3<0等价于RN>1, 此时方程 (3.1) 存在唯一的正根; 当B3>0时可推出RA<1, 此时B2>0成立, 此时方程 (3.1) 没有正根. 又因为L∗N>0等价于RνN>1, 则当且仅当RνN>1,RN>1时, 平衡点E6=(Sa6,Ia6,P6,Sc6,Ic6,L∗N,F∗N,IN6,0,0,0)存在.
下面计算只存在血食蝇时模型的地方病平衡点, 令LN=FN=IN=0. 系统 (2.2) 的简化系统为
从系统 (3.2) 可以看出当RνH>1时, 在平衡点处,LH7=L∗H=ΛH+μHαH(RνH−1),FH7=F∗H=ΛHdHL∗H. 记m1=βPaP+βHaIHSa+Ia,m2=βaHIaSa+Ia, 此时 (3.2) 式的平衡点满足下列方程
此时
从而
将方程 (3.5) 代入方程 (3.4) 中有
下面仅考虑m1≠0的情况. 方程 (3.6) 两端同时除以m1可得
当δa=0时, 定义
显然, 当m_{1}\in[0,+\infty)时,g(m_{1})单调递减, 则当且仅当g(0)=\frac{\beta_{Pa}\eta_{c}\Lambda_{a}}{\xi\mu_{c}d_{a}}+\frac{\beta_{Ha}F_{H}^{\ast}\beta_{aH}}{d_{H}\Lambda_{a}}>1,即R_{H}>1时, 方程 (3.8) 存在唯一的正根, 即系统 (3.2) 存在唯一的正平衡点. 因此, 当且仅当R_{\nu H}>1,R_{H}>1时, 平衡点E_{7}=(S_{a_{7}},I_{a_{7}},P_{7},S_{c_{7}},I_{c_{7}},0,0,0,L_{H}^{\ast},F_{H}^{\ast},I_{H_{7}})存在.
考虑一般情况, 即\delta_{a}\geq0, 整理方程 (3.7) 可以得到
其中
显然C_{1}>0,C_{4}>0\Leftrightarrow R_{H}<1. 定义H(z)=z^{3}+\frac{C_{2}}{C_{1}}z^{2}+\frac{C_{3}}{C_{1}}z+\frac{C_{4}}{C_{1}},z_{1}=\frac{-C_{2}+\sqrt{C_{2}^{2}-3C_{1}C_{3}}}{3C_{1}}, 根据文献[20], 可以得到以下定理.
定理 3.1 假设\delta_{a}\geq0,R_{\nu H}>1, 则下面的结论成立
(i) 若R_{H}>1, 系统 (2.2) 至少存在一个形如(\tilde{S}_{a},\tilde{I}_{a},\tilde{P},\tilde{S}_{c},\tilde{I}_{c},0,0,0,L_{H}^{\ast},F_{H}^{\ast},\tilde{I}_{H})的平衡点.
(ii) 若R_{H}\leq1且\triangle=C_{2}^{2}-3C_{1}C_{3}<0, 系统 (2.2) 不存在形如(\tilde{S}_{a},\tilde{I}_{a},\tilde{P},\tilde{S}_{c},\tilde{I}_{c},0,0,0,L_{H}^{\ast},F_{H}^{\ast},\tilde{I}_{H})的平衡点.
(iii) 若R_{H}\leq1, 则当且仅当z_{1}>0且H(z_{1})\leq0时, 系统 (2.2) 存在形如(\tilde{S}_{a},\tilde{I}_{a},\tilde{P},\tilde{S}_{c},\tilde{I}_{c},0,0,0,L_{H}^{\ast},F_{H}^{\ast},\tilde{I}_{H})的平衡点.
最后, 计算系统 (2.2) 的正平衡点. 设n_{1}=\beta_{Pa}P+\frac{\beta_{Ha}I_{H}}{S_{a}+I_{a}},n_{2}=\frac{\beta_{cN}I_{c}}{S_{c}+I_{c}},n_{3}=\frac{\beta_{aH}I_{a}}{S_{a}+I_{a}}, 讨论过程与上述方法相同, 可知当\delta_{a}=0时, 若R_{\nu N}>1,R_{\nu H}>1,R_{0}>1均成立, 则系统 (2.2) 存在唯一的正平衡点E_{8}=(S_{a_{8}},I_{a_{8}},P_{8},S_{c_{8}},I_{c_{8}},L_{N}^{\ast},F_{N}^{\ast},I_{N_{8}},L_{H}^{\ast},F_{H}^{\ast},I_{H_{8}}). 对于一般情况, 即\delta_{a}\geq0, 定义
其中
依据文献[21]定义p=\frac{D_{2}}{D_{1}}, q=\frac{D_{3}}{D_{1}}, u=\frac{D_{4}}{D_{1}}, v=\frac{D_{5}}{D_{1}}, p_{1}=\frac{q}{2}-\frac{3}{16}p^{2}, q_{1}=\frac{p^{3}}{32}-\frac{pq}{8}+u, D=\left(\frac{q_{1}}{2}\right)^{2}+\left(\frac{p_{1}}{3}\right)^{3}, \sigma=\frac{-1+\sqrt{3}i}{2},y_{1}=\sqrt [3]{-\frac{q_{1}}{2}+\sqrt{D}}+\sqrt[3]{-\frac{q_{1}}{2}-\sqrt{D}}, y_{2}=\sqrt[3]{-\frac{q_{1}}{2}+\sqrt{D}}\sigma+\sqrt[3]{-\frac{q_{1}}{2}-\sqrt{D}}\sigma^{2}, y_{3}=\sqrt[3]{-\frac{q_{1}}{2}+\sqrt{D}}\sigma^{2}+\sqrt[3]{-\frac{q_{1}}{2}-\sqrt{D}}\sigma, w_{i}=y_{i}-\frac{3p}{4},i=1,2,3. 易知v>0等价于R_{0}<1. 由文献[21]可以得到以下定理.
定理 3.2 假设R_{\nu N}>1,R_{\nu H}>1, 则下面结论成立
(i) 如果R_{0}>1, 系统 (2.2) 至少存在一个正平衡点.
(ii) 如果R_{0}\leq1且D\geq0, 当且仅当w_{1}>0,Y(w_{1})<0时系统 (2.2) 存在正平衡点.
(iii) 如果R_{0}\leq1且D<0, 系统 (2.2) 存在正平衡点当且仅当至少存在一个w^{\ast}\in\{w_{1},w_{2},w_{3}\}, 使得w^{\ast}>0且Y(w^{\ast})\leq0.
4 稳定性分析
4.1 局部渐近稳定性
下面分析系统 (2.2) 平衡点的局部渐近稳定性. 由于平衡点E_{6},E_{7}和E_{8}的稳定性不易分析, 因此只考虑\delta_{a}=0的情况.
定理 4.1 平衡点E_{1}=(S_{a}^{0},0,0,S_{c}^{0},0,0,0,0,0,0,0), E_{2}=(S_{a_{2}},I_{a_{2}},P_{2},S_{c_{2}},I_{c_{2}},0,0,0, 0, 0,0), E_{4}=(S_{a}^{0},0,0,S_{c}^{0},0,L_{N}^{\ast},F_{N}^{\ast},0,0,0,0)和E_{6}=(S_{a_{6}},I_{a_{6}},P_{6},S_{c_{6}},I_{c_{6}},L_{N}^{\ast},F_{N}^{\ast},I_{N_{6}},0, 0,0)局部渐近稳定的条件如下
(i) 当R_{A}<1,R_{\nu N}<1且R_{\nu H}<1时, 平衡点E_{1}局部渐近稳定; 若R_{A}>1或R_{\nu N}>1或R_{\nu H}>1, 平衡点E_{1}不稳定.
(ii) 当R_{A}>1时, 平衡点E_{2}存在. 若R_{\nu N}<1,R_{\nu H}<1, 平衡点E_{2}局部渐近稳定; 若R_{\nu N}>1或R_{\nu H}>1, 平衡点E_{2}不稳定.
(iii) 当R_{\nu N}>1时, 平衡点E_{4}存在. 若R_{\nu H}<1,R_{N}<1, 平衡点E_{4}局部渐近稳定; 若R_{\nu H}>1或R_{N}>1, 平衡点E_{4}不稳定.
(iv) 当R_{N}>1且R_{\nu N}>1时, 平衡点E_{6}存在. 当\delta_{a}=0时, 若R_{\nu H}<1, 平衡点E_{6}局部渐近稳定; 若R_{\nu H}>1, 平衡点E_{6}不稳定.
证 对于平衡点E_{1}, 其雅可比矩阵的形式为
其中
由分块矩阵的性质可知,J(E_{1})的特征值由\tilde{J}_{11}和\tilde{J}_{22}的特征值组成.\tilde{J}_{11}的特征方程为
其中I_{5\times5}是五阶单位矩阵, 且
-M_{1}的顺序主子式为\tilde{\triangle}_{1}=d_{a}+\delta_{a},\tilde{\triangle}_{2}=\xi(d_{a}+\delta_{a}),\tilde{\triangle}_{3}=(d_{a}+\delta_{a})\xi\mu_{c}(1-R_{A}),\tilde{\triangle}_{4}=\tilde{\triangle}_{3}(\mu_{N}+\Lambda_{N}),\tilde{\triangle}_{5}=\tilde{\triangle}_{3}d_{N}(\mu_{N}+\Lambda_{N})(1-R_{\nu N}), 当且仅当R_{A}<1,R_{\nu N}<1成立时,-M_{1}的顺序主子式均大于零. 又因为-M_{1}的主对角线元素均大于零, 非主对角线元素非正, 则-M_{1}是 M-矩阵. 由文献[22]可知, 若R_{A}<1,R_{\nu N}<1成立,-M_{1}的所有特征根均具有正实部, 即M_{1}的所有特征根均具有负实部. 由于方程 (4.1) 的另外两个特征根为-\mu_{c}和-d_{a}, 则当R_{A}<1,R_{\nu N}<1时,\tilde{J}_{11}的所有特征根均具有负实部.
\tilde{J}_{22}的特征方程为
显然, 当且仅当d_{H}(\mu_{H}+\Lambda_{H})-\Lambda_{H}b_{H}>0即R_{\nu H}<1时,\tilde{J}_{22}的所有特征根均具有负实部.
综上所述, 当且仅当R_{A}<1,R_{\nu N}<1且R_{\nu H}<1时, 平衡点E_{1}是局部渐近稳定的.
对于平衡点E_{2}和E_{4}, 其局部渐近稳定性的证明方法与E_{1}相同, 证明过程不再重复. 由于平衡点E_{6}的稳定性不易证明, 因此在证明稳定性时只考虑\delta_{a}=0的情况. 利用同样的思想, 可以得到: 当平衡点E_{6}存在时, 若R_{\nu H}<1, 则平衡点E_{6}是局部渐近稳定的. 证毕.
定理 4.2 平衡点E_{3}=(S_{a}^{0},0,0,S_{c}^{0},0,0,0,0,L_{H}^{\ast},F_{H}^{\ast},0), E_{5}=(S_{a}^{0},0,0,S_{c}^{0},0,L_{N}^{\ast},F_{N}^{\ast},0, L_{H}^{\ast},F_{H}^{\ast},0), E_{7}=(S_{a_{7}},I_{a_{7}},P_{7},S_{c_{7}},I_{c_{7}},0,0,0,L_{H}^{\ast},F_{H}^{\ast},I_{H_{7}}) 和 E_{8}=(S_{a_{8}},I_{a_{8}},P_{8},S_{c_{8}},I_{c_{8}},L_{N}^{\ast}, F_{N}^{\ast},I_{N_{8}},L_{H}^{\ast},F_{H}^{\ast},I_{H_{8}})局部渐近稳定的条件如下
(i) 当R_{\nu H}>1时, 平衡点E_{3}存在. 若R_{\nu N}<1,R_{A}<1, 平衡点E_{3}局部渐近稳定; 若R_{\nu N}>1或R_{A}>1, 平衡点E_{3}不稳定.
(ii) 当R_{\nu N}>1且R_{\nu H}>1时, 系统 (2.2) 的无病平衡点E_{5}存在. 若R_{0}<1, 平衡点E_{5}局部渐近稳定; 若R_{0}>1, 平衡点E_{5}不稳定.
(iii) 假设\delta_{a}=0, 当R_{\nu H}>1且R_{H}>1时, 平衡点E_{7}存在. 若R_{\nu N}<1, 平衡点E_{7}局部渐近稳定; 若R_{\nu N}>1, 平衡点E_{7}不稳定.
(iv) 假设\delta_{a}=0, 当R_{\nu N}>1,R_{\nu H}>1且R_{0}>1时, 系统 (2.2) 的地方病平衡点E_{8}存在, 此时平衡点E_{8}局部渐近稳定.
证 对于平衡点E_{3}, 它的特征方程为
其中I_{9\times9}是九阶单位矩阵, 且
其中,a_{99}=\mu_{H}+\Lambda_{H}+2\alpha_{H}L_{H}^{\ast}. -M_{2}的顺序主子式为\bar{\triangle}_{1}=d_{a}+\delta_{a}, \bar{\triangle}_{2}=\xi(d_{a}+\delta_{a}), \bar{\triangle}_{3}=\xi\mu_{c}(d_{a}+\delta_{a})(1-R_{A}), \bar{\triangle}_{4}=\bar{\triangle}_{3}(\mu_{N}+\Lambda_{N}), \bar{\triangle}_{5}=\bar{\triangle}_{3}d_{N}(\mu_{N}+\Lambda_{N})(1-R_{\nu N}), \bar{\triangle}_{6}=\bar{\triangle}_{5}d_{N}, \bar{\triangle}_{7}=\bar{\triangle}_{6}(\mu_{H}+\Lambda_{H}+2\alpha_{H}L_{H}^{\ast}), \bar{\triangle}_{8}=\bar{\triangle}_{6}d_{H}(\mu_{H}+\Lambda_{H})(R_{\nu H}-1), \bar{\triangle}_{9}=\bar{\triangle}_{8}d_{H}-\frac{\beta_{Ha}\beta_{aH}F_{H}^{\ast}\xi\mu_{c}d_{N}}{S_{a}^{0}}d_{N}(\mu_{N}+\Lambda_{N})(R_{\nu N}-1)d_{H}(\mu_{H}+\Lambda_{H})(R_{\nu H}-1), 当且仅当R_{A}<1,R_{\nu N}<1且R_{\nu H}>1成立时,-M_{2}的顺序主子式均大于零. 又因为-M_{2}的主对角线元素均大于零, 非主对角线元素非正, 则-M_{2}是 M-矩阵. 由文献[22]可知, 若R_{A}<1,R_{\nu N}<1且R_{\nu H}>1成立, 则-M_{2}的所有特征根均具有正实部, 即M_{2}的所有特征根均具有负实部. 由于方程 (4.3) 的另外两个特征根为-\mu_{c}和-d_{a}, 则当平衡点E_{3}存在时, 若R_{A}<1且R_{\nu N}<1, 平衡点E_{3}局部渐近稳定.
类似地, 可以证明该定理中的结论 (ii), (iii) 和 (iv). 证毕.
4.2 全局渐近稳定性
定理 4.3 平衡点E_{1}=(S_{a}^{0},0,0,S_{c}^{0},0,0,0,0,0,0,0)和E_{2}=(S_{a_{2}},I_{a_{2}},P_{2},S_{c_{2}},I_{c_{2}},0,0,0,0,0,0)全局渐近稳定的条件如下
(i) 若R_{\nu N}<1,R_{\nu H}<1且R_{A}<1, 则平衡点E_{1}在\Omega内部全局渐近稳定.
(ii) 若R_{\nu N}<1,R_{\nu H}<1且R_{A}>1, 则平衡点E_{2}在\Omega内部全局渐近稳定.
证 (i) 由引理 2.1 可知, 若R_{\nu N}<1,R_{\nu H}<1, 则当t\rightarrow\infty时有(L_{N}(t),F_{N}(t))\rightarrow(0,0),(L_{H}(t),F_{H}(t))\rightarrow(0,0), 此时I_{N}(t)\rightarrow0,I_{H}(t)\rightarrow0, 系统 (2.2) 的极限系统为
由定理 4.1 可知, 当R_{A}<1,R_{\nu N}<1且R_{\nu H}<1时, 平衡点E_{1}局部渐近稳定. 由于R_{A}=\frac{\beta_{Pa}\eta_{c}\Lambda_{a}}{d_{a}\mu_{c}\xi}<1, 则存在充分小的\varepsilon>0, 使得\frac{\beta_{Pa}\eta_{c}(\frac{\Lambda_{a}}{d_{a}}+\varepsilon)}{\mu_{c}\xi}<1. 由式 (2.3) 可知\mathop{\limsup}\limits_{t\rightarrow\infty}N_{a}(t)\leq \frac{\Lambda_{a}}{d_{a}}, 则存在充分大的t>0, 使得N_{a}(t)\leq\frac{\Lambda_{a}}{d_{a}}+\varepsilon. 构造 Lyapunov 函数
沿系统 (4.4) 的轨线求V_{1}的全导数可得
当R_{A}<1时V'_{1}\leq0, 且V'_{1}=0当且仅当P=0成立, 此时有I_{c}=0, 从而当t\rightarrow\infty时, 有S_{a}(t)\rightarrow S_{a}^{0},I_{a}(t)\rightarrow 0,S_{c}(t)\rightarrow S_{c}^{0}. 因此,V'_{1}=0的唯一紧不变子集为\{\tilde{E}_{1}=(S_{a}^{0},0,0,S_{c}^{0},0)\}. 由 LaSalle 不变集原理[23]可知\tilde{E}_{1}=(S_{a}^{0},0,0,S_{c}^{0},0)在\mathbb{R}_{+}^{5}上是全局渐近稳定的. 利用内链传递集理论[24]可知E_{1}=(S_{a}^{0},0,0,S_{c}^{0},0,0,0,0,0,0,0)全局渐近稳定.
(ii) 由 (i) 可以看出, 当R_{\nu N}<1,R_{\nu H}<1时, 系统 (4.4) 为系统 (2.2) 的极限系统. 为了简化系统, 引入新的变量, 令
系统 (4.4) 变为如下形式
定义 Lyapunov 函数V_{2}: Int(\mathbb{R}_{+}^{5})\mapsto \mathbb{R}
沿系统 (4.5) 的轨线求V_{2}的全导数可得
当且仅当x_{1}=x_{2}=x_{3}=x_{5}=1时V'_{2}=0. 结合系统 (4.5) 得当t\rightarrow\infty时x_{4}\rightarrow1. 由 LaSalle 不变集原理[23]可知,S_{a}(t)\rightarrow S_{a_{2}},I_{a}(t)\rightarrow I_{a_{2}},P(t)\rightarrow P_{2},S_{c}(t)\rightarrow S_{c_{2}},I_{c}(t)\rightarrow I_{c_{2}}. 利用内链传递集理论[24]可知E_{2}=(S_{a_{2}},I_{a_{2}},P_{2},S_{c_{2}},I_{c_{2}},0,0,0,0,0,0)全局渐近稳定. 证毕.
定理 4.4 平衡点E_{3}=(S_{a}^{0},0,0,S_{c}^{0},0,0,0,0,L_{H}^{\ast},F_{H}^{\ast},0), E_{4}=(S_{a}^{0},0,0,S_{c}^{0},0,L_{N}^{\ast},F_{N}^{\ast},0,0, 0,0) 和E_{5}=(S_{a}^{0},0,0,S_{c}^{0},0,L_{N}^{\ast},F_{N}^{\ast},0,L_{H}^{\ast},F_{H}^{\ast},0)全局渐近稳定的条件如下
(i) 若R_{\nu H}>1,R_{\nu N}<1且
则平衡点E_{3}在\Omega内部是全局渐近稳定的.
(ii) 若R_{\nu N}>1,R_{\nu H}<1,R_{N}<1且
则平衡点E_{4}在\Omega内部是全局渐近稳定的.
(iii) 若R_{\nu N}>1,R_{\nu H}>1,R_{0}<1且
则平衡点E_{5}在\Omega内部是全局渐近稳定的.
证 由定理 4.2 可知, 若R_{\nu H}>1,R_{\nu N}<1且R_{A}<1成立, 平衡E_{3}是局部渐近稳定的. 由引理 2.1 可得, 若R_{\nu N}<1,R_{\nu H}>1, 则当t\rightarrow\infty时有(L_{N}(t),F_{N}(t))\rightarrow(0,0),(L_{H}(t),F_{H}(t))\rightarrow(L_{H}^{\ast},F_{H}^{\ast}), 从而当t\rightarrow\infty时可得I_{N}(t)\rightarrow0, 此时, 系统 (2.2) 的极限系统为
由系统 (4.6) 的前两个方程可得:\Lambda_{a}-(d_{a}+\delta_{a})N_{a}(t)\leq N'_{a}(t)\leq\Lambda_{a}-d_{a}N_{a}(t), 因此
其中\phi_{1}=\frac{\Lambda_{a}}{d_{a}+\delta_{a}},S_{a}^{0}=\frac{\Lambda_{a}}{d_{a}}. 当t充分大时有
考虑系统 (4.7) 的辅助系统
定义
显然,-J的顺序主子式为\check{\triangle}_{1}=d_{a}+\delta_{a},\check{\triangle}_{2}=\xi(d_{a}+\delta_{a}), \check{\triangle}_{3}=\xi\mu_{c}(d_{a}+\delta_{a})(1-R_{A}), \check{\triangle}_{4}=\xi\mu_{c}(d_{a}+\delta_{a})d_{H}\left(1-R_{A}-\left(\frac{d_{a}+\delta_{a}}{d_{a}}\right)^{2}\frac{\beta_{Ha}\beta_{aH}F_{H}^{\ast}}{d_{H}S_{a}^{0}(d_{a}+\delta_{a})}\right). 当R_{A}+\left(\frac{d_{a}+\delta_{a}}{d_{a}}\right)^{2}\frac{\beta_{Ha}\beta_{aH}F_{H}^{\ast}}{d_{H}S_{a}^{0}(d_{a}+\delta_{a})}<1时, \check{\triangle}_{3}>0,\check{\triangle}_{4}>0. 定义
易知R_{A}+\left(\frac{d_{a}+\delta_{a}}{d_{a}}\right)^{2}\frac{\beta_{Ha}\beta_{aH}F_{H}^{\ast}}{d_{H}S_{a}^{0}(d_{a}+\delta_{a})}<1等价于R_{H}<\tilde{R}_{c}, 则当R_{H}<\tilde{R}_{c},-J的顺序主子式均大于零. 又因为-J的主对角线元素大于零, 非主对角线元素非正, 则-J为 M-矩阵, 此时系统 (\ref{4.8}) 的平衡点(0,0,0,0)局部渐近稳定, 由于系统 (4.8) 是一个线性系统, 则平衡点(0,0,0,0)在\mathbb{R}_{+}^{4}上全局渐近稳定. 由比较原理可知, 当t\rightarrow\infty时有(I_{a},P,I_{c},I_{H})\rightarrow(0,0,0,0), 从而当t\rightarrow\infty时, 有S_{a}(t)\rightarrow S_{a}^{0},S_{c}(t)\rightarrow S_{c}^{0}. 因此, 当R_{\nu H}>1,R_{\nu N}<1且R_{H}<\tilde{R}_{c}时, 平衡点E_{3}全局渐近稳定.
采用类似的方法可以证明结论 (ii) 和 (iii). 证毕.
推论 4.1 假设\delta_{a}=0, 此时\tilde{R}_{c}=1, 从而若R_{\nu H}>1,R_{\nu N}<1且R_{H}<1成立, 平衡点E_{3}在\Omega内部是全局渐近稳定的.
推论 4.2 假设\delta_{a}=0, 此时\hat{R}_{c}=1, 从而若R_{N}<1,R_{\nu N}>1且R_{\nu H}<1成立, 平衡点E_{4}在\Omega内部是全局渐近稳定的.
推论 4.3 假设\delta_{a}=0, 此时R_{A}+\left(\frac{d_{a}+\delta_{a}}{d_{a}}\right)\frac{\beta_{Pa}\beta_{cN}\eta_{N}F_{N}^{\ast}}{\xi d_{a}d_{N}}+\left(\frac{d_{a}+\delta_{a}}{d_{a}}\right)^{2}\frac{\beta_{aH}\beta_{Ha}d_{a}F_{H}^{\ast}}{\Lambda_{a}d_{H}(d_{a}+\delta_{a})}<1等价于R_{0}<1, 从而若R_{\nu N}>1,R_{\nu H}>1且R_{0}<1, 平衡点E_{5}在\Omega内部是全局渐近稳定的.
由于系统 (2.2) 的维数比较高, 很难确定平衡点E_{6},E_{7}和E_{8}的全局稳定性, 因此, 仅在\delta_{a}=0的情况下对其全局稳定性进行研究.
定理 4.5 假设\delta_{a}=0, 则平衡点E_{6}=(S_{a_{6}},I_{a_{6}},P_{6},S_{c_{6}},I_{c_{6}},L_{N}^{\ast},F_{N}^{\ast},I_{N_{6}},0,0,0), E_{7}=(S_{a_{7}},I_{a_{7}},P_{7},S_{c_{7}},I_{c_{7}},0,0,0,L_{H}^{\ast},F_{H}^{\ast},I_{H_{7}}) 和E_{8}=(S_{a_{8}},I_{a_{8}},P_{8},S_{c_{8}},I_{c_{8}},L_{N}^{\ast},F_{N}^{\ast},I_{N_{8}},L_{H}^{\ast},F_{H}^{\ast}, I_{H_{8}})全局渐近稳定的条件如下
(i) 若R_{\nu N}>1,R_{N}>1且R_{\nu H}<1, 则平衡点E_{6}在\Omega内部是全局渐近稳定的.
(ii) 若R_{\nu H}>1,R_{H}>1且R_{\nu N}<1, 则平衡点E_{7}在\Omega内部是全局渐近稳定的.
(iii) 若R_{\nu N}>1,R_{\nu H}>1且R_{0}>1, 则平衡点E_{8}在\Omega内部是全局渐近稳定的.
证 由定理 4.1 可知当\delta_{a}=0时, 若R_{N}>1,R_{\nu N}>1且R_{\nu H}<1, 平衡点E_{6}是局部渐近稳定的. 由引理 2.1 可知, 若R_{\nu N}>1,R_{\nu H}<1, 则当t\rightarrow\infty时有(L_{N}(t),F_{N}(t))\rightarrow(L_{N}^{\ast},F_{N}^{\ast}),(L_{H}(t),F_{H}(t))\rightarrow(0,0), 从而当t\rightarrow\infty时可得I_{H}(t)\rightarrow 0. 由系统 (2.2) 得
显然, 当t\rightarrow\infty时,(N_{a}(t),N_{c}(t))\rightarrow(S_{a}^{0},S_{c}^{0}), 系统 (4.9) 的唯一正平衡点(S_{a}^{0},S_{c}^{0})是全局渐近稳定的, 此时系统 (2.2) 的极限系统为
设K:=[S_{a}^{0}]\times[\frac{\eta_{c}S_{c}^{0}+\eta_{N}F_{N}^{\ast}}{\xi}]\times[S_{c}^{0}]\times[F_{N}^{\ast}], 则\omega((I_{a}(0),P(0),I_{c}(0),I_{H}(0)))\subset K, 其中\omega((I_{a}(0),P(0),I_{c}(0),I_{H}(0)))是(I_{a}(0),P(0),I_{c}(0),I_{H}(0))\in \mathbb{R}_{+}^{4}对于系统 (4.10) 的解半流的\omega极限集. 设
则f:\mathbb{R}_{+}^{4}\rightarrow \mathbb{R}^{4}是一个连续可微映射, 此外f具有以下性质
(1)f在K上是合作的, 且对任一v\in K,Df(v)=\left(\frac{\partial f_{i}}{\partial v_{j}}\right)_{1\leq i,j\leq4}是不可约的;
(2) 对于所有的v\in K,f(0)=0且当v_{i}=0时,f_{i}(v)\geq0,i=1,2,3,4;
(3) 对任意的\rho\in(0,1),(v_{1},v_{2},v_{3},v_{4})\inIntK, 有
类似可得f_{2}(\rho v_{1},\rho v_{2},\rho v_{3},\rho v_{4})=\rho f_{2}(v_{1},v_{2},v_{3},v_{4}), f_{3}(\rho v_{1},\rho v_{2},\rho v_{3},\rho v_{4})=\rho f_{3}(v_{1},v_{2},v_{3},v_{4}), f_{4}(\rho v_{1},\rho v_{2},\rho v_{3},\rho v_{4})>\rho f_{4}(v_{1},v_{2},v_{3},v_{4}), 则f在K上是严格次线性的.
由于
则
定义Df(0)的谱界为
易知当R_{A}+\frac{\beta_{Pa}\beta_{cN}F_{N}^{\ast}\eta_{N}}{\xi d_{a}d_{N}}>1即R_{N}>1时, det(Df(0))<0,Df(0)至少存在一个正的特征值, 从而s(Df(0))>0. 由文献[16]可知, 若R_{N}>1, 系统 (4.10) 的正平衡点(I_{a_{6}},P_{6},I_{c_{6}},I_{H_{6}})在\mathbb{R}_{+}^{4}\setminus\{(0,0,0,0)\}中是全局渐近稳定的. 从而, 当t\rightarrow\infty时,
因此, 当\delta_{a}=0时, 若R_{N}>1,R_{\nu N}>1且R_{\nu H}<1, 平衡点E_{6}全局渐近稳定.
类似可以证明平衡点E_{7}和平衡点E_{8}的全局稳定性. 证毕.
4.3 持久性
定理 4.5 中讨论了\delta_{a}=0时正平衡点的全局稳定性, 对于\delta_{a}\geq0的一般情况, 正平衡点的全局稳定性不易得到, 下面的定理给出了疾病的持久性.
若R_{\nu N}>1,R_{\nu H}>1,R_{0}>1, 则存在一个\epsilon>0, 使得对于系统 (2.2) 中I_{a}(0)>0,P(0)>0,I_{c}(0)>0,I_{N}(0)>0,I_{H}(0)>0的所有解满足
5 数值模拟
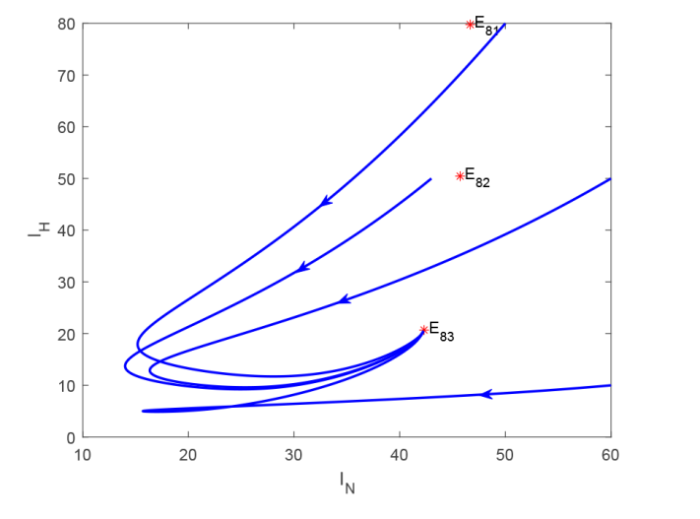
上面在证明平衡点{{E}_{7}}和{{E}_{8}}存在性时发现, 当染病食草动物的因病死亡率{{\delta }_{a}}\ge 0时不止存在一个此类型的平衡点, 并且在证明其稳定性时, 仅仅分析了{{\delta }_{a}}=0时的特殊情况, 不够全面, 下面以{{E}_{8}}为例, 给出系统 (2.2) 相应的参数值, 通过数值模拟分析其稳定性情况. 令\Lambda_{a}=1, \beta_{Pa}=0.001, \beta_{Ha}=0.02, d_{a}=\frac{1}{800}, \delta_{a}=\frac{1}{10}, \eta_{c}=0.002, \eta_{N}=0.01, \xi=\frac{1}{10}, \mu_{c}=0.05, b_{N}=3, \mu_{N}=0.1, \Lambda_{N}=\frac{1}{16}, \alpha_{N}=0.05, b_{H}=3, \mu_{H}=0.1, \Lambda_{H}=\frac{1}{16}, \alpha_{H}=0.05, \beta_{cN}=0.01, \beta_{aH}=0.05, d_{N}=\frac{1}{30}, d_{H}=\frac{1}{30}. 此时R_{vN}=34.6154>1, R_{vH}=34.6154>1, R_{0}=2.4>1, 系统 (2.2) 存在三个正平衡点, 分别为:E_{81}=(13.132,9.7144,5.0607,0.3283,19.6717,109.25,204.8438,46.6725,109.25,204.8438,79.7719), E_{82}=(33.9559,9.4573,4.9543,0.8489,19.1511,109.25,204.8438,45.713,109.25,204.8438,50.4506), E_{83}=(105.8201,8.5701,4.5781,2.6455,17.3545,109.25,204.8438,42.3103,109.25,204.8438,20.6947). 由图 1 可以看出, 正平衡点E_{83}是渐近稳定的, 正平衡点E_{81}和E_{82}是不稳定的.
图1
为了降低炭疽导致的食草动物的发病率和死亡率, 探讨不同因素对炭疽传播的影响是非常重要的. 为此, 讨论一些参数与R_{0}的关系, 以及控制炭疽传播的可能措施.
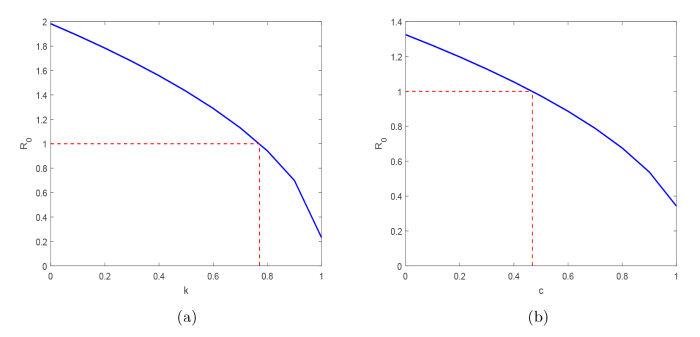
由于尸食性蝇和血食性蝇均能传播炭疽, 因此叮咬率对炭疽的传播起着非常重要的作用. 对于尸食性蝇, 为了模拟防止染病尸体被尸食性蝇叮咬导致炭疽传播的效果, 在模型中使用(1-k)\beta_{cN}代替\beta_{cN}, 其中\Lambda_{a}=0.1,\beta_{Pa}=0.01,\beta_{Ha}=0.2,\eta_{N}=0.001,\mu_{c}=0.1,b_{N}=0.3,b_{H}=0.3,\beta_{cN}=0.1,\beta_{aH}=0.015,其余参数值与图 1 中的参数值相同. 图 2(a) 显示了干预措施k与基本再生数R_{0}的关系, 其中R_{0}是关于k的递减函数. 因此, 可以通过清除动物尸体, 减少尸食性蝇的食物来源等方法对炭疽传播进行控制.
对于血食性蝇, 为了模拟易感动物被血食性蝇叮咬导致炭疽传播的效果, 在模型中使用(1-c)\beta_{Ha}代替\beta_{Ha}, 其中\beta_{Pa}=0.005,\beta_{Ha}=0.2,\delta_{a}=\frac{1}{7},\beta_{cN}=0.005,\beta_{aH}=0.2, 其余参数值与图 2 中的参数值相同. 图 2(b) 显示了干预措施c与基本再生数R_{0}的关系, 其中R_{0}是关于c的递减函数. 因此, 可以在苍蝇成熟之前的阶段清理苍蝇的繁殖地点, 杀死幼年的苍蝇, 并且对成年苍蝇使用杀虫剂, 减少苍蝇数量, 降低血食性蝇感染炭疽的风险, 从而减少血食性蝇对易感动物的叮咬, 降低炭疽的传播速率.
图2
6 结论
该文研究了尸食性蝇、血食性蝇两类媒介对动物种群中炭疽传播的影响. 通过定义各类再生数, 对模型平衡点的存在性及稳定性进行了分析, 且分析了疾病的持久性, 并通过数值模拟研究了染病尸体对尸食性蝇的传染率\beta_{cN}、染病的血食性蝇对易感动物的传染率\beta_{Ha}两者对模型基本再生数R_{0}的影响. 结果表明: 通过清除动物尸体能够减少尸食性蝇的食物来源, 从而降低模型的基本再生数, 使用杀虫剂减少血食性蝇数量, 能够降低血食性蝇对易感动物的叮咬, 从而降低模型的基本再生数, 两种措施对炭疽传播均起到了抑制作用.
参考文献
Spores and soil from six sides: Interdisciplinarity and the environmental biology of anthrax (Bacillus anthracis)
DOI:10.1111/brv.2018.93.issue-4 URL [本文引用: 1]
Anthrax undervalued zoonosis
DOI:10.1016/j.vetmic.2009.08.016 URL [本文引用: 1]
A mathematical model of an anthrax epizoötic in the Kruger National Park
DOI:10.1016/0307-904X(81)90034-2 URL [本文引用: 1]
Anthrax epizootic and migration: Persistence or extinction
DOI:10.1016/j.mbs.2012.10.004
PMID:23137874
[本文引用: 1]

In this paper, we use an extension of the deterministic mathematical model of an anthrax epizootic of Hahn and Furniss to study the effects of anthrax transmission, carcass ingestion, carcass induced environmental contamination, and migration rates on the persistence or extinction of animal populations. We compute the basic reproduction number R(0) for the anthrax epizootic model with and without taking into account animal migration. We obtained conditions for an anthrax enzootic region. We demonstrate that decreasing the levels of carcass ingestion by removal of carcases in game reserves, for example, may not always lead to a reduction in the population of animals infected with anthrax. However, increasing levels of carcass induced environmental contamination rates in an enzootic anthrax region can result in the catastrophic extinction of a persistent animal population.Copyright © 2012 Elsevier Inc. All rights reserved.
A mathematical model of anthrax transmission in animal populations
DOI:10.1007/s11538-016-0238-1
PMID:28035484
[本文引用: 1]
A general mathematical model of anthrax (caused by Bacillus anthracis) transmission is formulated that includes live animals, infected carcasses and spores in the environment. The basic reproduction number [Formula: see text] is calculated, and existence of a unique endemic equilibrium is established for [Formula: see text] above the threshold value 1. Using data from the literature, elasticity indices for [Formula: see text] and type reproduction numbers are computed to quantify anthrax control measures. Including only herbivorous animals, anthrax is eradicated if [Formula: see text]. For these animals, oscillatory solutions arising from Hopf bifurcations are numerically shown to exist for certain parameter values with [Formula: see text] and to have periodicity as observed from anthrax data. Including carnivores and assuming no disease-related death, anthrax again goes extinct below the threshold. Local stability of the endemic equilibrium is established above the threshold; thus, periodic solutions are not possible for these populations. It is shown numerically that oscillations in spore growth may drive oscillations in animal populations; however, the total number of infected animals remains about the same as with constant spore growth.
Species identification of adult African blowflies (diptera: Calliphoridae) of forensic importance
DOI:10.1007/s00414-017-1654-y
PMID:28849264
[本文引用: 1]
Necrophagous blowflies can provide an excellent source of evidence for forensic entomologists and are also relevant to problems in public health, medicine, and animal health. However, access to useful information about these blowflies is constrained by the need to correctly identify the flies, and the poor availability of reliable, accessible identification tools is a serious obstacle to the development of forensic entomology in the majority of African countries. In response to this need, a high-quality key to the adults of all species of forensically relevant blowflies of Africa has been prepared, drawing on high-quality entomological materials and modern focus-stacking photomicroscopy. This new key can be easily applied by investigators inexperienced in the taxonomy of blowflies and is made available through a highly accessible online platform. Problematic diagnostic characters used in previous keys are discussed.
Analysis of the Bulgarian tabanid fauna with regard to its potential for epidemiological involvement
The necrophagous fly anthrax transmission pathway: Empirical and genetic evidence from wildlife epizootics
DOI:10.1089/vbz.2013.1538 URL [本文引用: 1]
Human anthrax outbreak associated with livestock exposure: Georgia, 2012
Anthrax toxin
DOI:10.1146/cellbio.2003.19.issue-1 URL [本文引用: 1]
Transmission of pathogens by Stomoxys flies (Diptera, Muscidae): A review
DOI:10.1051/parasite/2013026
PMID:23985165
[本文引用: 1]
Stomoxys flies are mechanical vectors of pathogens present in the blood and skin of their animal hosts, especially livestock, but occasionally humans. In livestock, their direct effects are disturbance, skin lesions, reduction of food intake, stress, blood loss, and a global immunosuppressive effect. They also induce the gathering of animals for mutual protection; meanwhile they favor development of pathogens in the hosts and their transmission. Their indirect effect is the mechanical transmission of pathogens. In case of interrupted feeding, Stomoxys can re-start their blood meal on another host. When injecting saliva prior to blood-sucking, they can inoculate some infected blood remaining on their mouthparts. Beside this immediate transmission, it was observed that Stomoxys may keep some blood in their crop, which offers a friendly environment for pathogens that could be regurgitated during the next blood meal; thus a delayed transmission by Stomoxys seems possible. Such a mechanism has a considerable epidemiological impact since it allows inter-herd transmission of pathogens. Equine infectious anemia, African swine fever, West Nile, and Rift Valley viruses are known to be transmitted by Stomoxys, while others are suspected. Rickettsia (Anaplasma, Coxiella), other bacteria and parasites (Trypanosoma spp., Besnoitia spp.) are also transmitted by Stomoxys. Finally, Stomoxys was also found to act as an intermediate host of the helminth Habronema microstoma and may be involved in the transmission of some Onchocerca and Dirofilaria species. Being cosmopolite, Stomoxys calcitrans might have a worldwide and greater impact than previously thought on animal and human pathogen transmission. © F. Baldacchino et al., published by EDP Sciences, 2013.
Confirmation of Bacillus anthracis from flesh-eating flies collected during a West Texas anthrax season
This case study confirms the interaction between necrophilic flies and white-tailed deer, Odocoileus virginianus, during an anthrax outbreak in West Texas (summer 2005). Bacillus anthracis was identified by culture and PCR from one of eight pooled fly collections from deer carcasses on a deer ranch with a well-documented history of anthrax. These results provide the first known isolation of B. anthracis from flesh-eating flies associated with a wildlife anthrax outbreak in North America and are discussed in the context of wildlife ecology and anthrax epizootics.
Dynamical analysis and control strategies in modeling anthrax
DOI:10.1007/s40314-015-0297-1 URL [本文引用: 1]
Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems
Global asymptotic behavior in some cooperative systems of functional differential equations
Modelling malaria control by introduction of larvivorous fish
DOI:10.1007/s11538-011-9628-6
PMID:21347816
[本文引用: 2]
Malaria creates serious health and economic problems which call for integrated management strategies to disrupt interactions among mosquitoes, the parasite and humans. In order to reduce the intensity of malaria transmission, malaria vector control may be implemented to protect individuals against infective mosquito bites. As a sustainable larval control method, the use of larvivorous fish is promoted in some circumstances. To evaluate the potential impacts of this biological control measure on malaria transmission, we propose and investigate a mathematical model describing the linked dynamics between the host-vector interaction and the predator-prey interaction. The model, which consists of five ordinary differential equations, is rigorously analysed via theories and methods of dynamical systems. We derive four biologically plausible and insightful quantities (reproduction numbers) that completely determine the community composition. Our results suggest that the introduction of larvivorous fish can, in principle, have important consequences for malaria dynamics, but also indicate that this would require strong predators on larval mosquitoes. Integrated strategies of malaria control are analysed to demonstrate the biological application of our developed theory.
The construction of next-generation matrices for compartmental epidemic models
DOI:10.1098/rsif.2009.0386
PMID:19892718
[本文引用: 1]
The basic reproduction number (0) is arguably the most important quantity in infectious disease epidemiology. The next-generation matrix (NGM) is the natural basis for the definition and calculation of (0) where finitely many different categories of individuals are recognized. We clear up confusion that has been around in the literature concerning the construction of this matrix, specifically for the most frequently used so-called compartmental models. We present a detailed easy recipe for the construction of the NGM from basic ingredients derived directly from the specifications of the model. We show that two related matrices exist which we define to be the NGM with large domain and the NGM with small domain. The three matrices together reflect the range of possibilities encountered in the literature for the characterization of (0). We show how they are connected and how their construction follows from the basic model ingredients, and establish that they have the same non-zero eigenvalues, the largest of which is the basic reproduction number (0). Although we present formal recipes based on linear algebra, we encourage the construction of the NGM by way of direct epidemiological reasoning, using the clear interpretation of the elements of the NGM and of the model ingredients. We present a selection of examples as a practical guide to our methods. In the appendix we present an elementary but complete proof that (0) defined as the dominant eigenvalue of the NGM for compartmental systems and the Malthusian parameter r, the real-time exponential growth rate in the early phase of an outbreak, are connected by the properties that (0) > 1 if and only if r > 0, and (0) = 1 if and only if r = 0.
Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission
DOI:10.1016/S0025-5564(02)00108-6 URL [本文引用: 1]
On the zeros of a third degree exponential polynomial with applications to a delayed model for the control of testosterone secretion
DOI:10.1093/imammb/18.1.41 URL [本文引用: 1]
On the zeros of a fourth degree exponential polynomial with applications to a neural network model with delays
DOI:10.1016/j.chaos.2005.01.019 URL [本文引用: 2]
The Stability of Dynamical Systems
Chain transitivity, attractivity, and strong repellors for semidynamical systems
DOI:10.1023/A:1009044515567 URL [本文引用: 2]
Impact of visitors and hospital staff on nosocomial transmission and spread to community
DOI:10.1016/j.jtbi.2014.04.003 URL [本文引用: 1]
Spread of disease with transport-related infection and entry screening
An SIQS model is proposed to study the effect of transport-related infection and entry screening. If the basic reproduction number is below unity, the disease free equilibrium is locally asymptotically stable. There exists an endemic equilibrium which is locally asymptotically stable if the reproduction number is larger than unity. It is shown that the disease is endemic in the sense of permanence if and only if the endemic equilibrium exists. Entry screening is shown to be helpful for disease eradication since it can always have the possibility to eradicate the disease led by transport-related infection and furthermore have the possibility to eradicate disease even when the disease is endemic in both isolated cities.

{kind=link}
{kind=link}
{kind=link}
{kind=link}