

1 引言
种群生态学是研究控制生物种群量与其栖境关系的一门学科[1].法国数学家Poincare创立了微分方程定性理论[2].国内外的生物数学家利用微分方程定性理论在种群生态学领域取得了比较多的研究成果.由于生态系统的复杂和多样性,为了更准确的描述和刻画现实世界,科学家们将时滞引入到微分方程中对生态系统进行研究.捕食者与猎物的关系是自然界中生物种群相互作用的基本作用之一,关于它的各种情况的研究,很多学者做了大量的工作,也取得了不小的进展.如张天然在模型中加入了猎物性别因素[3],张丽红考虑到年龄结构的影响[4],这些因素的加入弥补了以前捕食者-猎物系统对种群内部相异性思考不足的情况,使得捕食者-猎物系统与现实的相符程度比之从前更进一步.
本文主要研究基于瓢虫、棉蚜和蚂蚁的捕食者-猎物-共生者系统,考虑到昆虫的时滞的影响更为显著,因此本文研究了具有时滞的捕食者-猎物-共生者系统,通过对模型的分析,了解瓢虫、棉蚜和蚂蚁之间的制约关系,为棉田的生态调节提供一定的理论依据.
2 预备知识
正性
定义2.1[9] 系统
系统(2.1)的解的正性是指当初值
持久性
定义2.2[10] 系统(2.1)持久性是指存在一个紧集
可以这样理解持久性,即生态系统在一定边界范围内保持恒定或维持某一特定状态的持续时间.
平衡点局部稳定性的判定
设种群、群落或生态系统的动力系统模型由微分方程表示,用向量形式写为
其中状态向量
特征值的分析是稳定性分析的关键,对于非线性微分方程,可先将其线性化,列出其特征方程,然后进行讨论.
定理2.1[11] 平衡点
反之,如果特征方程具有正实部的特征值,那么
的一切均有负实部的充要条件是
定理2.2[10] 关于方程
此时第三个条件也可写为
其中
3 模型的建立
根据文献[5]中的模型,本文主要建立并研究系统
该系统的各个参数生物学意义如下:
4 模型的分析
本部分主要对模型的正性,持久性和局部稳定性进行分析.系统(3.1)满足初始条件(4.1)和(4.2).
4.1 模型的正性分析
定理4.1 如果系统满足初始条件,则当
证 由系统(3.1)可知
对于第三个式子,首先假设当
即在
即在
综上,当初始条件(4.1)和(4.2)成立,所以当
4.2 模型的持久性分析
记
并记
定理4.2 若系统(3.1),满足条件(A1)和(A2),则系统(3.1)具有持久性.
证 因为
利用文献[10]中的方法,可知存在
因为
于是存在
因为
于是存在
根据文献[11],可以得出,若条件(A1)成立,存在
因为
由于
于是存在
综上所述,若条件(A2)成立,则存在
其中
4.3 平衡点的局部稳定性
系统(3.1)的平衡点有
其中
下面分别讨论这些平衡点的局部稳定性,对系统(3.1)在各个平衡点进行线性化,从而得出该方程平衡点
将
定理4.3 平衡点
定理4.4 若条件(A3):
证 平衡点
整理得
其中
下面分析
把(4.5)变成以下的形式
当条件(A3)条件成立时,该平衡点
记
定理4.5 在
证 对正平衡点
相应的特征方程整理得
设
此时,取
令
根据Hurwitz判据,有
满足条件(A4);
满足条件(A5);
满足条件(A6).
由此可知在
从生物学上看,
5 数值模拟
下面对系统(3.1)所做的平衡点稳定性分析进行数值模拟,对于情况较为复杂的平衡点进行数值分析.
(1) 平衡点
若条件(A3)成立,则平衡点
图 1
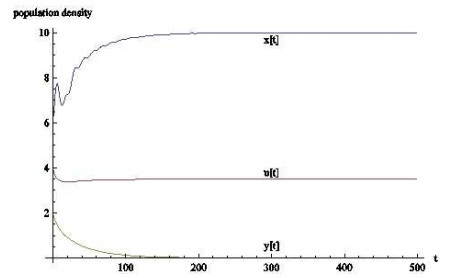
生物学上看,当瓢虫和棉蚜的时滞忽略不计时,棉蚜和蚂蚁可以共存并最终保持稳定,而瓢虫的数量会一直减少,甚至会消失.
(2) 平衡点
对于正平衡点
图 2
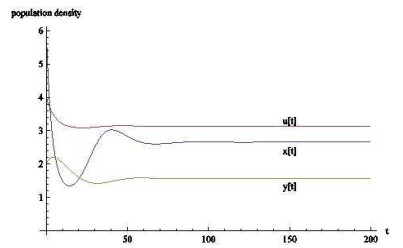
图 3

图 4

图 5
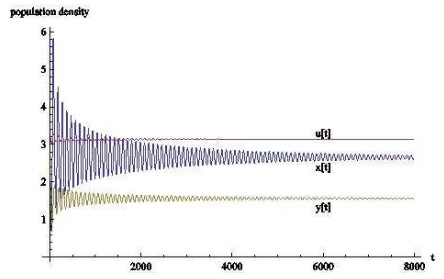
图 6

图 7
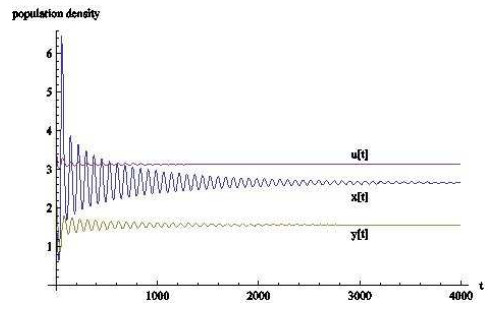
图 8

图 9
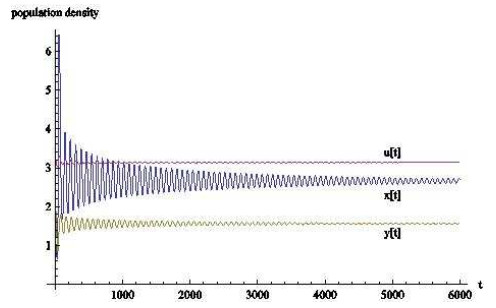
图 10
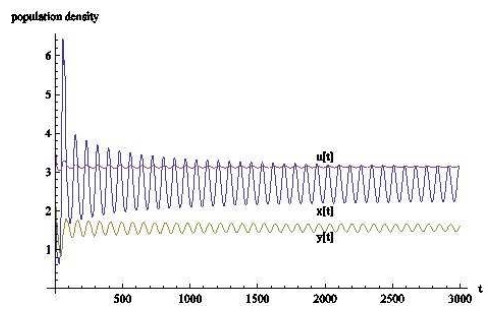
从数值模拟的结果来看,时滞对于基于瓢虫,棉蚜和蚂蚁的捕食者-猎物-共生者系统有相当大的影响.当时滞的作用忽略不计时,三种群最终会稳定在一个固定数值上;
从生物学上看,当瓢虫和棉蚜的发育历期不断增长时,三者虽然会共存但数量会不停变化,可以这样认为,发育历期影响着自身种群的发展,并间接影响着其余种群的发展.
图 11
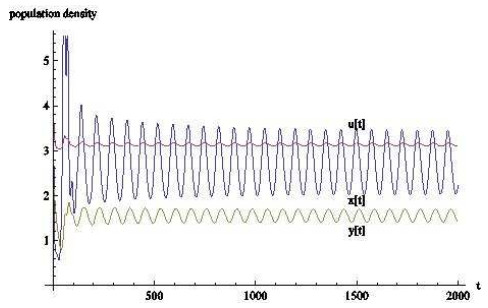
6 结果与讨论
对于本文所建立的捕食者-猎物-共生系统的模型,猎物和捕食者的时滞会对平衡点的稳定性起到重要作用,当时滞超过临界值时,数值模拟表明,系统会发生由稳定到周期波动的改变,说明捕食者和猎物的成长时间对系统会有相当重要的影响,从生物学上看,随着瓢虫和棉蚜的时滞的增长,即当发育历期的影响越来越大时,三者的关系会出现周期波动,这也更符合现实世界的情况.但由于棉蚜和瓢虫的发育历期会受温度、地域、气候等相关因素的影响,这里只采取了比较普遍的情况,所以可能在精确度上会有所影响.但同时结果表明,只有天敌可以部分抑制棉蚜的数量,但可能不足以控制害虫.如何有效控制害虫将是下一步要做的工作.
这对现实中的棉田有害生物的治理有所启发,判断棉蚜和瓢虫的发育历期,根据发育历期的长短预估将来发展的趋势,以此来设计相应的治理方案,如根据波动的情况来采取不同的治理方法,尤其要小心有剧烈波动时所产生的蚜虫大爆发的情况.
参考文献
随机配对种群的两性模型的动力学性态
DOI:10.3969/j.issn.1001-9626.2008.04.001 [本文引用: 1]
The dynamics of random mating two-sex population
DOI:10.3969/j.issn.1001-9626.2008.04.001 [本文引用: 1]
Hopf bifurcation in symmetric configuration of predator-prey mutualiststems
DOI:10.1016/j.na.2009.02.127 [本文引用: 2]
Turing pattern formation in a predator-prey-mutualist system
DOI:10.1016/j.nonrwa.2011.05.022 [本文引用: 1]
Dynamic behaviors of a discrete periodic predator-prey-mutualist system
Premanence of the periodic predator-prey-mutualist system
DOI:10.1186/s13662-015-0654-9 [本文引用: 2]
Permanence and stability of a predator-prey system with stage structure for predator
DOI:10.1016/j.cam.2005.12.035 [本文引用: 1]
带时滞的食蚜蝇-蚜虫年龄结构的捕食模型
A Predator-prey model for the age structure of syrphidae and aphids with time delay

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}