Inhibition of α-Synuclein Aggregation by the Interaction Between Protein Disulfide Isomerase and α-Synuclein
PEI Yun-shan1,2, ZHANG Cai1,2, LIU Xiao-li1, CHENG Kai1, ZHANG Ze-ting,1, LI Cong-gang1
1. CAS Key Laboratory of Magnetic Resonance in Biological Systems, State Key Laboratory of Magnetic Resonance and Atomic and Molecular Physics, Innovation Academy for Precision Measurement Science and Technology, Chinese Academy of Sciences, Wuhan 430071, China
2. University of Chinese Academy of Sciences, Beijing 100049, China
Abnormally misfolded and aggregated α-synuclein (αsyn) is the hallmark of Parkinson's disease (PD). Molecular chaperone protein disulfide isomerase (PDI) has been shown to interact with αsyn and inhibit its aggregation in vitro, but the mechanism for the recognition of αsyn by PDI is not yet clear. Herein, we used nuclear magnetic resonance (NMR) spectroscopy to identify that human PDI b'xa' bound with the N-terminal domain of αsyn, and thioflavin T (ThT) fluorescence assay revealed that b'xa' domain of PDI significantly inhibited αsyn aggregation. Furthermore, by using NMR titration, we observed that PDI bound to αsyn mainly through its hydrophobic cavity of the b' domain. Based on these findings, a docking model of PDI binding with αsyn was established and a possible mechanism of how PDI inhibits αsyn aggregation was proposed. Our work provides experimental evidences for understanding the inhibitory role of PDI in αsyn aggregation.
Keywords:nuclear magnetic resonance (NMR)
;
α-synuclein (αsyn)
;
protein disulfide isomerase
;
interaction
;
fibrillation
PEI Yun-shan. Inhibition of α-Synuclein Aggregation by the Interaction Between Protein Disulfide Isomerase and α-Synuclein. Chinese Journal of Magnetic Resonance[J], 2022, 39(4): 381-392 doi:10.11938/cjmr20222974
Fig.1
The b'xa' domain of PDI interacts with N-terminal domain of αsyn. (a) Model for PDI (red) primary sequence, PDI ab domain (pink) and PDI b'xa' domain (blue) are shown underneath. (b) Surface crystal structure of human PDI (PDB ID: 4EKZ). (c) Protein purities analyzed by SDS-PAGE. M, Marker; lane 1, αsyn1-60; lane 2, αsyn; lane 3, PDI ab; lane 4, PDI b'xa'; lane 5, PDI. (d) and (e) Spectral overlay of αsyn in the absence (black) and presence of equivalent amount of PDI (red) and PDI b'xa' (blue). Residues showing significant signal attenuation or shifts were marked. (f) Residue-resolved attenuation (I/I0) of αsyn in the presence of PDI (red), PDI ab (pink) and PDI b'xa' (blue). (g) Hydrophobic residues of αsyn predicted by OMH method, the binding and hydrophobic areas are colored gray
Fig.2
The effect on αsyn aggregation of PDI, PDI ab and PDI b'xa' detected by ThT experiment. (a) Aggregations kinetics of αsyn in the absence (black) and presence of equimolar PDI ab (pink), PDI b'xa' (blue) and PDI (red). (b) The fitted half-time (t1/2) values of aggregation and their statistical analysis. ns: not significant, ****: p < 0.0001; the dots represent the values obtained from four parallel experiments
Fig.3
NMR results showing the interaction between PDI b'xa' with the N-terminal domain of αsyn. Overlaid 1H-15N TROSY spectra of PDI b'xa' (red) with (blue) αsyn 1-19 (a), αsyn 30-42 (d) or αsyn 1-60 (g). E239 were selected as representative residues for global chemical shift perturbations (CSPs) during NMR titration. Residue-resolved CSPs of PDI b'xa' during titrations with αsyn 1-19(b), 30-42(e) and 1-60 (h). KD values were fitted from curves of residues with significant CSPs, as a function of αsyn 1-19(c), αsyn 30-42(f), and αsyn 1-60 (i) concentration. Residues showing significant CSPs were marked in spectra and mapped red on the crystal structure surface of PDI b'xa'. Threshold level was indicated as 'mean+standard deviation'. Disperse residues were verified and removed by the threshold of 'mean +2×standard deviation'
Fig.4
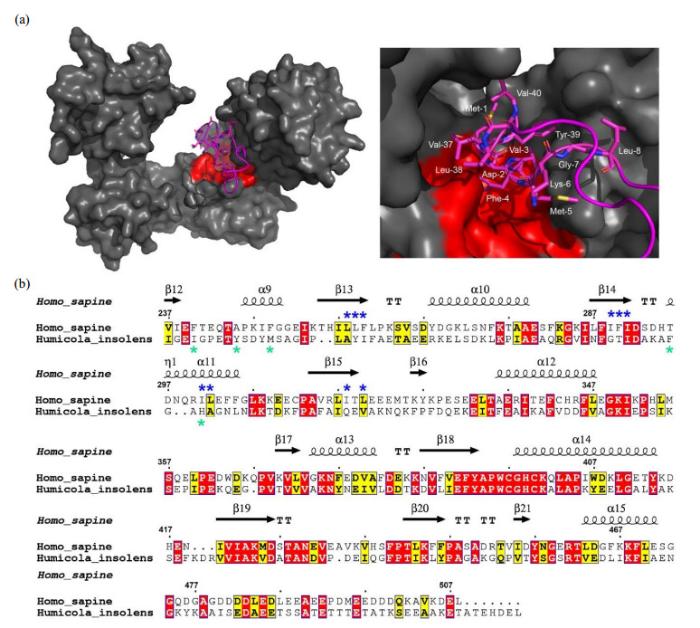
Hydrophobic binding mode of PDI with the N-terminal domain of αsyn. (a) Docking model of PDI binding αsyn1-40 (left panel) and a local zoom view of binding area (right panel). The relevant residues of αsyn1-40 involving in binding PDI were depicted as magenta sticks. HDOCK experiments were performed on the web server (http://hdock.phys.hust.edu.cn/) by uploading human PDI crystal structure, amino acid sequences of αsyn1-40 and their binding sites identified by NMR titration. Optimal docking results were plotted with PyMol software, PDI molecular surface was colored gray and its hydrophobic pocket was marked in red, the magenta cartoon represented αsyn1-40 structure. (b) Alignment of amino acid sequence of human PDI b'xa' from HiPDI b'xa'. Sequence identity and similarity were calculated by Clustalw[52] and ESPript 3.0[53]. Similar amino acids were showed as black letters in yellow boxes and identical amino acids were showed as white letters in red boxes. The secondary structural domains were indicated on the top of the sequences. Blue and green asterisks represented the key hydrophobic residues of αsyn interacting with human PDI and HiPDI, respectively
A precipitating role for truncated alpha-synuclein and the proteasome in alpha-synuclein aggregation - Implications for pathogenesis of Parkinson disease
Interaction of alpha-synuclein with divalent metal ions reveals key differences: A link between structure, binding specificity and fibrillation enhancement
Interaction of alpha-synuclein with divalent metal ions reveals key differences: a link between structure, binding specificity and fibrillation enhancement
Both the isomerase and chaperone activities of protein disulfide isomerase are required for the reactivation of reduced and denatured acidic phospholipase A2
Quantitative analysis of multisite protein-ligand interactions by NMR: binding of intrinsically disordered p53 transactivation subdomains with the TAZ2 domain of CBP
A precipitating role for truncated alpha-synuclein and the proteasome in alpha-synuclein aggregation - Implications for pathogenesis of Parkinson disease
0
2005
New insights into cellular α-synuclein homeostasis in health and disease
0
2016
α-Synuclein in Lewy bodies
0
1997
α-synuclein – regulator of exocytosis, endocytosis, or both?
0
2017
The process of Lewy body formation, rather than simply α-synuclein fibrillization, is one of the major drivers of neurodegeneration
0
2020
Lewy pathology in Parkinson's disease consists of crowded organelles and lipid membranes
Interaction of alpha-synuclein with divalent metal ions reveals key differences: A link between structure, binding specificity and fibrillation enhancement
Ca2+ binding to alpha-synuclein regulates ligand binding and oligomerization
0
2001
Interaction of alpha-synuclein with divalent metal ions reveals key differences: a link between structure, binding specificity and fibrillation enhancement
Both the isomerase and chaperone activities of protein disulfide isomerase are required for the reactivation of reduced and denatured acidic phospholipase A2
Quantitative analysis of multisite protein-ligand interactions by NMR: binding of intrinsically disordered p53 transactivation subdomains with the TAZ2 domain of CBP
1
2012
... 结合常数KD由(2)式[42]拟合得到: ...
Alpha-synuclein stepwise aggregation reveals features of an early onset mutation in Parkinson's disease
... 结合NMR滴定实验结果和还原型人源PDI晶体结构(PDB ID: 4EKZ),我们使用HDOCK服务[51]对接得到PDI与αsyn1-40的结合模型如图 4(a)所示.最佳对接模型中,PDI主要通过b'上的疏水空腔结合αsyn N端区域.对接模型的局部放大图显示,αsyn1-40蛋白以1~8号和37~40号的疏水氨基酸残基将PDI b'上的疏水口袋完全占据,与我们提出的PDI结合αsyn的方式为疏水相互作用相吻合.Yagi-Utsumi等[32]的研究结果显示,结合人源αsyn31-41多肽时,HiPDI b'xa'以氧化态形式参与反应,还原态则不能结合底物.而本研究显示,人源PDI及PDI b'xa'在还原条件下均能有效结合αsyn并抑制其聚集.然而,由于人源氧化态PDI及PDI b'xa'较还原条件下更易发生多聚[24],导致无法获得高质量的NMR谱图,因此本研究未对氧化态PDI与αsyn的相互作用展开研究.由于HiPDI比人源PDI b'结构域的疏水空腔面积小,且序列比对结果显示HiPDI与人源PDI的序列相似度不高,疏水口袋b'区域(237~347号氨基酸残基)序列相似度仅为31.5%.因此结合αsyn多肽时,氧化态HiPDI参与相互作用的关键残基分布与人源PDI也有所差别[图 4(b)].但是HiPDI仍通过b'结构域疏水残基参与αsyn多肽的相互作用[32],与本研究中人源PDI通过b'结构域的疏水空腔结合αsyn的结果相似(图 4),表明两种不同来源的PDI都以疏水相互作用结合αsyn底物.PDI与<i>α</i>syn N端的疏水结合模式. (a) PDI结合<i>α</i>syn 1-40多肽的对接模型(左)及其结合区域的局部放大图(右),HDOCK对接实验在网站服务器(http://hdock.phys.hust.edu.cn/)上运行,上传人源PDI晶体结构,<i>α</i>syn1-40序列及NMR滴定鉴定的结合位点,最佳对接结果使用PyMol软件作图,PDI表面结构呈灰色,疏水口袋以红色标出,<i>α</i>syn1-40卡通结构呈品红色,右图中参与结合的<i>α</i>syn残基以棒状模式显示并标出.(b)人源PDI b'xa'与<i>Hi</i>PDI b'xa'的氨基酸序列比对.序列一致性和相似性由Clustalw<sup>[<xref ref-type="bibr" rid="b52">52</xref>]</sup>和ESPript 3.0<sup>[<xref ref-type="bibr" rid="b53">53</xref>]</sup>分析所得,相似氨基酸(黑色字母填充黄色方框)以及相同氨基酸(白色字母填充红色方框)分别被标出,顶部标注人源PDI b'xa'二级结构区域,蓝色和绿色星号分别表示<i>α</i>syn结合人源PDI和<i>Hi</i>PDI的关键疏水氨基酸残基Hydrophobic binding mode of PDI with the N-terminal domain of <i>α</i>syn. (a) Docking model of PDI binding <i>α</i>syn1-40 (left panel) and a local zoom view of binding area (right panel). The relevant residues of <i>α</i>syn1-40 involving in binding PDI were depicted as magenta sticks. HDOCK experiments were performed on the web server (http://hdock.phys.hust.edu.cn/) by uploading human PDI crystal structure, amino acid sequences of <i>α</i>syn1-40 and their binding sites identified by NMR titration. Optimal docking results were plotted with PyMol software, PDI molecular surface was colored gray and its hydrophobic pocket was marked in red, the magenta cartoon represented <i>α</i>syn1-40 structure. (b) Alignment of amino acid sequence of human PDI b'xa' from <i>Hi</i>PDI b'xa'. Sequence identity and similarity were calculated by Clustalw<sup>[<xref ref-type="bibr" rid="b52">52</xref>]</sup> and ESPript 3.0<sup>[<xref ref-type="bibr" rid="b53">53</xref>]</sup>. Similar amino acids were showed as black letters in yellow boxes and identical amino acids were showed as white letters in red boxes. The secondary structural domains were indicated on the top of the sequences. Blue and green asterisks represented the key hydrophobic residues of <i>α</i>syn interacting with human PDI and <i>Hi</i>PDI, respectivelyFig.4
... [52] and ESPript 3.0[53]. Similar amino acids were showed as black letters in yellow boxes and identical amino acids were showed as white letters in red boxes. The secondary structural domains were indicated on the top of the sequences. Blue and green asterisks represented the key hydrophobic residues of αsyn interacting with human PDI and HiPDI, respectively Fig.4
Deciphering key features in protein structures with the new ENDscript server
2
2014
... 结合NMR滴定实验结果和还原型人源PDI晶体结构(PDB ID: 4EKZ),我们使用HDOCK服务[51]对接得到PDI与αsyn1-40的结合模型如图 4(a)所示.最佳对接模型中,PDI主要通过b'上的疏水空腔结合αsyn N端区域.对接模型的局部放大图显示,αsyn1-40蛋白以1~8号和37~40号的疏水氨基酸残基将PDI b'上的疏水口袋完全占据,与我们提出的PDI结合αsyn的方式为疏水相互作用相吻合.Yagi-Utsumi等[32]的研究结果显示,结合人源αsyn31-41多肽时,HiPDI b'xa'以氧化态形式参与反应,还原态则不能结合底物.而本研究显示,人源PDI及PDI b'xa'在还原条件下均能有效结合αsyn并抑制其聚集.然而,由于人源氧化态PDI及PDI b'xa'较还原条件下更易发生多聚[24],导致无法获得高质量的NMR谱图,因此本研究未对氧化态PDI与αsyn的相互作用展开研究.由于HiPDI比人源PDI b'结构域的疏水空腔面积小,且序列比对结果显示HiPDI与人源PDI的序列相似度不高,疏水口袋b'区域(237~347号氨基酸残基)序列相似度仅为31.5%.因此结合αsyn多肽时,氧化态HiPDI参与相互作用的关键残基分布与人源PDI也有所差别[图 4(b)].但是HiPDI仍通过b'结构域疏水残基参与αsyn多肽的相互作用[32],与本研究中人源PDI通过b'结构域的疏水空腔结合αsyn的结果相似(图 4),表明两种不同来源的PDI都以疏水相互作用结合αsyn底物.PDI与<i>α</i>syn N端的疏水结合模式. (a) PDI结合<i>α</i>syn 1-40多肽的对接模型(左)及其结合区域的局部放大图(右),HDOCK对接实验在网站服务器(http://hdock.phys.hust.edu.cn/)上运行,上传人源PDI晶体结构,<i>α</i>syn1-40序列及NMR滴定鉴定的结合位点,最佳对接结果使用PyMol软件作图,PDI表面结构呈灰色,疏水口袋以红色标出,<i>α</i>syn1-40卡通结构呈品红色,右图中参与结合的<i>α</i>syn残基以棒状模式显示并标出.(b)人源PDI b'xa'与<i>Hi</i>PDI b'xa'的氨基酸序列比对.序列一致性和相似性由Clustalw<sup>[<xref ref-type="bibr" rid="b52">52</xref>]</sup>和ESPript 3.0<sup>[<xref ref-type="bibr" rid="b53">53</xref>]</sup>分析所得,相似氨基酸(黑色字母填充黄色方框)以及相同氨基酸(白色字母填充红色方框)分别被标出,顶部标注人源PDI b'xa'二级结构区域,蓝色和绿色星号分别表示<i>α</i>syn结合人源PDI和<i>Hi</i>PDI的关键疏水氨基酸残基Hydrophobic binding mode of PDI with the N-terminal domain of <i>α</i>syn. (a) Docking model of PDI binding <i>α</i>syn1-40 (left panel) and a local zoom view of binding area (right panel). The relevant residues of <i>α</i>syn1-40 involving in binding PDI were depicted as magenta sticks. HDOCK experiments were performed on the web server (http://hdock.phys.hust.edu.cn/) by uploading human PDI crystal structure, amino acid sequences of <i>α</i>syn1-40 and their binding sites identified by NMR titration. Optimal docking results were plotted with PyMol software, PDI molecular surface was colored gray and its hydrophobic pocket was marked in red, the magenta cartoon represented <i>α</i>syn1-40 structure. (b) Alignment of amino acid sequence of human PDI b'xa' from <i>Hi</i>PDI b'xa'. Sequence identity and similarity were calculated by Clustalw<sup>[<xref ref-type="bibr" rid="b52">52</xref>]</sup> and ESPript 3.0<sup>[<xref ref-type="bibr" rid="b53">53</xref>]</sup>. Similar amino acids were showed as black letters in yellow boxes and identical amino acids were showed as white letters in red boxes. The secondary structural domains were indicated on the top of the sequences. Blue and green asterisks represented the key hydrophobic residues of <i>α</i>syn interacting with human PDI and <i>Hi</i>PDI, respectivelyFig.4
... [53]. Similar amino acids were showed as black letters in yellow boxes and identical amino acids were showed as white letters in red boxes. The secondary structural domains were indicated on the top of the sequences. Blue and green asterisks represented the key hydrophobic residues of αsyn interacting with human PDI and HiPDI, respectively Fig.4







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}